Nature |
您所在的位置:网站首页 › 能量atp会在哪些细胞器中产生 › Nature |
Nature
|
真核细胞的脂质主要在内质网 (endoplasmic reticulum, ER) 合成,其由内质网向其他有膜细胞器的传递主要通过细胞器之间的膜接触位点 (membrane contact site, MCS) 进行1 (图1)。在酵母中,ER-线粒体之间的MCS由ERMES (ER-mitochondira encounter structure) 介导2,该复合物由4个核心成分组成,包括Mmm1、Mdm12、Mdm34和Mdm102,其中Mmm1、Mdm12和Mdm34均含有保守的SMP (synaptotagmin-like mitochondrial lipid-binding protein) 结构域,该结构域存在于许多酵母和哺乳动物MCS脂质转移蛋白中3。虽然目前已有包括Mmm1和Mdm12在内的SMP结构域结的结构,SMP结构域如何在两个细胞器之间排列并介导细胞器间的脂质运输仍不清楚4。
图1. 有膜细胞器之间不同类型的MCS5 2023年5月10日,来自MRC的Wanda Kukulski和来自弗里堡大学的Stefano Vanni共同通讯,在Nature上在线发表了题为In situ architecture of the ER–mitochondria encounter structure的科研论文。该研究通过定量活细胞成像、冷冻电镜技术,确定了ERMES在Saccharomyces cerevisiae中的分子组成,此外,结合分子动力学模拟,研究者还建立了ERMES的分子模型,对ER和线粒体间的脂质转移途径进行了进一步的阐述和猜想。 利用定量活细胞荧光显微镜,研究者首先通过荧光强度对每个ERMES中含SMP结构域组分的数量和比例进行了探究,发现Mmm1、Mdm34以及Mdm12的荧光强度在整体上存在相关性 (图2)。在过去的体外实验中,Mmm1、Mdm34、Mdm12以等摩尔量形成稳定的复合物6,与本文所观察到的现象相一致。
图2. 通过荧光强度对每个MCS中ERMES组分的计量比进行确定 为进一步探究ER和线粒体MCS处ERMES的分子结构及其所发挥的生物学功能,研究者利用cryo-CLEM (correlative light and electron cryo-microscopy) 对Saccharomyces cerevisiae的ERMES MCS进行成像和数据采集 (图3)。
图3. 利用cryo-CLEM进行数据采集 数据分析结果显示,在ER和线粒体的MCS处,存在许多密集的桥梁状结构,这些结构跨越两个细胞器之间的间隙,整体长度为24.2 nm ± 4.76 nm (图4)。通过STA (subtomogram averaging),研究者得到的密度图能够吻合ER和线粒体之间的间隙,可能对应于ERMES复合物完整的细胞质部分。该密度图由三段大小和形状类似的结构组成,呈锯齿状排列,具备一定的灵活性,大体上可以以两种不同的长度存在。Mmm1锚定在ER膜上并于Mdm12相互作用,Mdm34与Mdm12以及线粒体外膜蛋白Mdm10相互作用6,提示在本文获得的密度图中,近ER一侧的蛋白组分极有可能为Mmm1,中部为Mdm12,而近线粒体一侧为Mdm34。这种组织方式可以适应不同的桥接长度,并且可以通过同时锚定两个细胞器膜来稳定地连接ER和线粒体。
图4. ER和线粒体之间的桥梁状结构 在过去的体外研究中,Mmm1和Mdm34的SMP结构域均可以发生同源二聚化6,因此研究者推测,这两者或许也可以介导ERMES桥梁的二聚化。为进一步确定这些桥梁结构彼此之间是否具备特定的排列方式,研究者进一步对ERMES在ER和线粒体外膜上的膜锚定点以及ERMES桥梁结构中心点的坐标进行分析 (图5)。虽然有一些相邻的桥梁结构可能彼此相关联,但大多数锚定点之间的距离数据表明相邻桥梁结构之间不存在显著的二聚化排列特征。
图5. 可能的二聚化模型以及坐标分析 此外,研究者发现,许多桥梁状的结构与膜并不垂直,而是存在一定的倾角。此外,在MCS中,ERMES桥梁呈不规则的簇状分布,虽然桥梁结构具备不同的长度,但长度相近的桥梁结构更容易在局部聚集成簇。在每个MCS中,ERMES桥梁的数量与MCS的面积成正比,表明ERMES桥的数量受到空间调节的限制 (图6)。
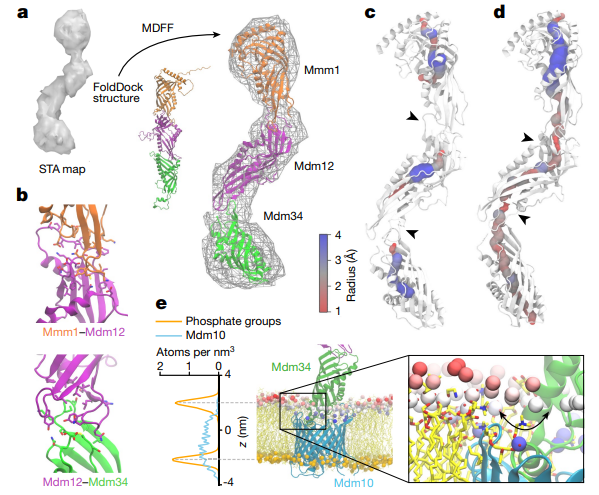
图6. ERMES在MCS上的空间分布 利用FoldDock预测得到的Mmm1-Mdm12异源二聚体以及Mdm12-Mdm34异源二聚体模型后,研究者将这些模型对接入前述STA密度图,发现Mmm1、Mdm12和Mdm34的SMP结构域头尾相接排列,前一个亚基的头端环状结构插入后一个亚基的裂隙中。与在细胞中观察到的锯齿状排列不同,FoldDock预测的异源三聚体复合物呈“线性”构象,通过分子动力学柔性拟合 (molecular dynamic flexible fitting, MDFF) 将预测的复合物结构做了进一步的调整。最终的模型显示,SMP结构域的空腔在Mdm12-Mdm34和Mmm1-Mdm12交界处变窄, RMES可能形成连续的脂质通道,但SMP结构域的这种排列方式可能会在亚基交界处限制脂质的转移。此外,研究者还构建了包含线粒体外膜蛋白Mdm10在内的完整ERMES模型,利用MDFF模拟,发现Mdm10在胞质侧表现出显著的疏水特性,可能与脂质分子的摄取和释放相关 (图7)。
图7. 完整ERMES复合物的分子模型 综上,本文通过解析ER和线粒体间介导脂质转移的ERMES的原位分子结构,揭示了ER和线粒体间的脂质转移通路,并为理解真核细胞中脂质转移的机制提供了基础。 -023-06050-3 参考文献 参考文献 1.Reinisch, K. M. & Prinz, W. A. Mechanisms of nonvesicular lipid transport. J. Cell Biol. 220, e202012058 (2021). 2.Kornmann, B. et al. An ER-mitochondria tethering complex revealed by a synthetic biology screen. Science 325, 477–481 (2009). 3.Kopec, K. O., Alva, V. & Lupas, A. N. Homology of SMP domains to the TULIP superfamily of lipid-binding proteins provides a structural basis for lipid exchange between ER and mitochondria. Bioinformatics 26, 1927–1931 (2010). 4. Schauder, C. M. et al. Structure of a lipid-bound extended synaptotagmin indicates a role in lipid transfer. Nature 510, 552–555 (2014). 5.Rossini, M. et al. Better to keep in touch: investigating inter-organelle cross-talk. FEBS J. 288: 740-755 (2021). 6.Jeong, H., Park, J., Jun, Y. & Lee, C. Crystal structures of Mmm1 and Mdm12–Mmm1 reveal mechanistic insight into phospholipid trafficking at ER–mitochondria contact sites. Proc. Natl Acad. Sci. USA 114, E9502–E9511 (2017). 供稿 | 王彤彤 审稿 | 丛野 责编 | 囡囡 排版 | 可洲 |






【本文地址】
今日新闻 |
推荐新闻 |