科学网 |
您所在的位置:网站首页 › 鳄鱼的进化过程图文解读 › 科学网 |
科学网
|
解读基因组的进化—从创新到“翻新”与“堆砌”
已有 24120 次阅读 2014-7-3 10:07 |系统分类:观点评述| 基因组, 有性生殖, C值悖论 没有基因就没有生命的繁衍,也就没有一切实体生命的存在。各种生物的基因组是一部部生物类群演化的史册,它记载了生物间的血脉关系与演化历程,昭示着生命的继承、承载与发展。因此,生命的演化映射于个体的基因组—种群的基因库—生命所有的基因集合这样一种极为复杂、多层次的遗传系统的相互关联与相互作用之中。 基因是控制生物性状的基本遗传单位,是携带有遗传信息的DNA序列,它是生命遗传系统的结构基础。大量的生物学家正在致力于以各种各样的(特别是模式)生物为对象通过基因及其功能的研究来解码生命,在微观的生命科学领域中取得了大量惊人的进展,但关于这些个别基因的信息细节不是本文的论述范畴。所谓基因组(genome)是指包含在一个生物体的一套染色体中完整的DNA序列(或全部遗传信息)。基因组包括基因和非编码DNA,它本质上是一套极为复杂的控制生命过程的操作系统,指挥生命活动(生长、发育、繁殖等)所有的遗传指令都匿藏于基因组之中。基因组还操控生命个体在各种生存环境中的复杂的响应与适应行为。 物种的进化必定是建立在基因组的进化之上,但两者是否完全等同或吻合?它们的演化轨迹是否完全一致?人们可否根据基因组的进化来透视物种的生态遗传学设计原理?
一、基因组大小—继承与随机的复杂化 1. 不太有章法的DNA—C值悖論 在每一种生物中,其单倍体基因组的DNA总量被称为C值 (C Value)。人们可能容易直观地人为,某种生物的基因组大小应该与其进化程度或复杂度正相关。但是大量的研究表明,物种的基因组大小与其在进化上所处地位的高低或复杂性没有绝对的相关性,这种现象称为C值悖論(C-ValueParadox)。 图1为各种类群生物的单倍体基因组大小,近似地,1 pg ≈十亿 bp (即1000 Mb or1 Gb),更准确地说,碱基对数=质量(pg)×0.978 × 109,或者质量(pg)=碱基对数1.022 × 10-9(Dolezel et al. 2003)。不难看出,基因组大小与进化程度并不怎么吻合,譬如肺鱼的基因组远大于哺乳动物,一些昆虫的基因组都能大于哺乳动物,而原生动物的基因组的变化范围最大,占据了所有生物最大基因组的位置(Gregory 2004)。 原生生物的基因组大小相差超过30万倍,动物基因组大小的差别也超过了3300倍,陆生植物相差约1000倍(Wikipedia)。 虽然一些类群的基因组变化巨大,但是从总的趋势来看,真核生物的基因组显著大于原核生物。虽然真核生物基因组大小的变异很大,但是从最小C值来看,一般进化程度越高的类群,群内最小C值越大。
图1 各种类群生物的单倍体基因组大小(“C-值”,pg)的范围(引自Gregory 2004)
2. 基因的进化—从朴素简洁到奢侈浪费? 随着人们对基因结构和功能认识的深入,人们将基因进一步区分为结构基因、调节基因和操纵基因:结构基因能为多肽链编码,调节基因能调节蛋白质的合成,而操纵基因为操纵结构基因的基因。 即便是结构基因也不一定全部由编码序列组成,有些在编码序列中间插入无编码作用的碱基序列,形成所谓断裂基因。一些基因的DNA序列包含两个区段:一个区段将被表达并存在于成熟的mRNA中,称为“外显子”,一个区段由虽然也同时被表达,但将在成熟mRNA中被删除,称为“内含子”。原核生物的基因序列一般是连续的,在一个基因的内部几乎不含“内含子”,而真核生物中绝大多数基因都是由不连续DNA序列组成的断裂基因。 基因组大小与编码蛋白的基因数目之间存在怎样的关系?Hou & Lin(2009)整理了各类生物的基因组及基因数目的资料,真核生物的基因组变化于373~3,175,581kbp之间,非真核生物(细菌、古菌、病毒、线粒体和叶绿体)的基因组变化于2.4~9950 kbp之间,因此真核生物的基因组远大于非真核生物的基因组。从图2不难看出,基因组与编码蛋白的基因数目之间存在显著的正相关关系,但是非真核生物和真核生物的直线回归方程的斜率不同,前者显著大于后者。由于每个基因组的编码蛋白的基因数一般与总基因数非常接近,用总基因数代替编码蛋白的基因数,类似与图2A的关系也完全成立。
图2 基因组大小与编码蛋白基因数目(A)或基因编码(构成基因的DNA部分)百分比(B)之间的关系(引自Hou & Lin 2009)
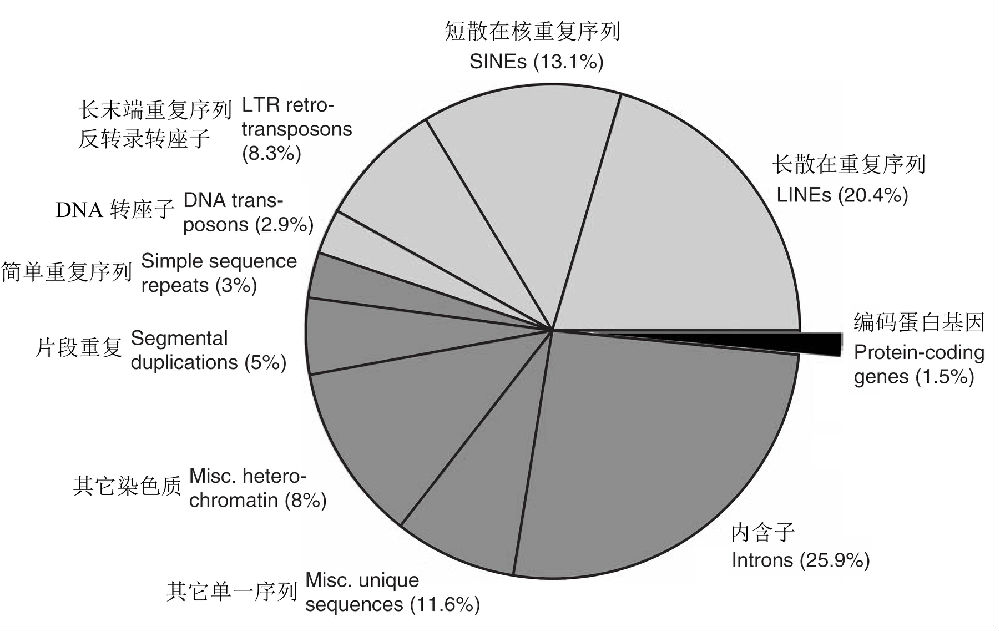
关于基因组大小与编码基因的DNA比例(%)之间的关系,在非真核生物中几乎就是一根平行的直线(除了一些细胞器的偏离值以外),而在真核生物中,则呈现明显的负相关关系(图2B)。具体来说,在真核生物中,随着基因组的增大,编码基因的DNA比例从81.6%下降到1.2%,而在非真核生物中,则保持较高的比例(97~47%),随基因组大小的变动幅度比真核生物显著地减小,只有细胞器基因组例外(Hou & Lin 2009)。仅从蛋白质生产的角度来看,原核生物的基因组似乎比真核生物更为高效。 人类基因组含有约30亿个DNA碱基对,曾估计可以形成10万个以上的基因,但事实上人类只有大约2~2.5万个基因,和老鼠相差无几(两者有99%的基因是相同的)。在人类基因组中,蛋白质编码序列(称为外显子)只占1.5%(图3),而其余均是不能编码蛋白质的序列! 看来,生物进化程度(等级)越高,非编码序列在基因组中的比重越大,譬如在微生物中,非编码区只占整个基因组序列的10%-20%,而在人类基因组中,这个比重高达98.5%!为什么高等生物的DNA会如此奢侈浪费?一些人称这些为“垃圾DNA”(junk DNA),认为这些序列大多数是演化的副产物,除了一些可能承载着重要的遗传信息外,多数可能已经没有什么作用了。
图3 人类基因组的组成:基因组中仅有约1.5%由严格的蛋白编码序列组成,而45%由各种类型的转座因子(浅灰色区域)构成,内含子占到26%,片段重复约占5%,数据源自International Human Genome Sequencing Consortium(2001)(Gregory 2005)
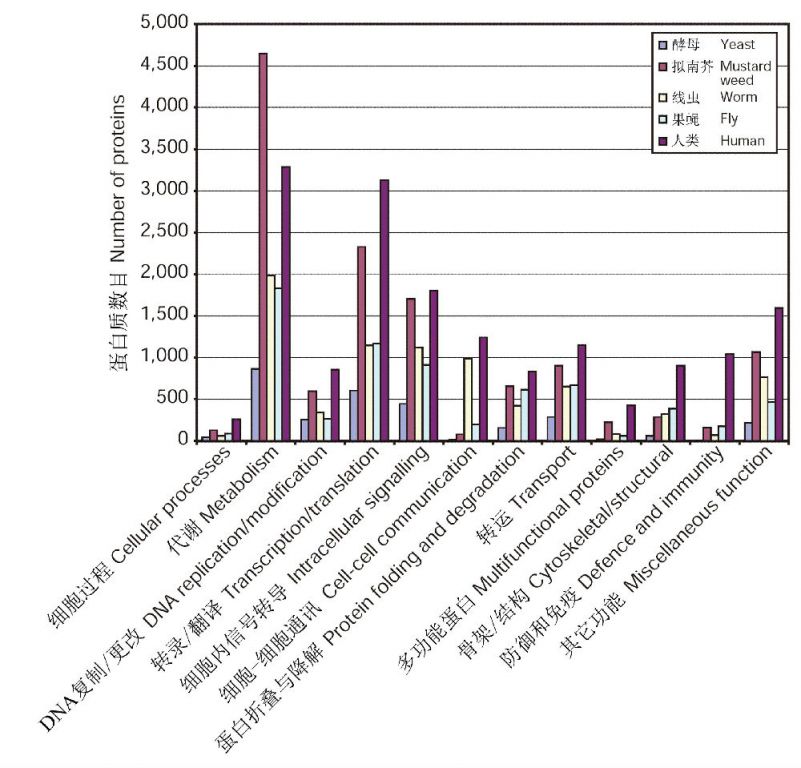
3. 基因的进化—继承与发展 蛋白质是基因的产物,是生命结构的基本材料。蛋白质的组成是基因组成的写照。从人类同源蛋白的分布可以看出,原核生物和真核生物共有的蛋白占到21%,动物和其它真核生物共有的蛋白占到32%,动物共有的蛋白占到24%,脊椎动物共有的蛋白占到22%(图4)。人类的很多基因起源自一些共同的祖先,但也包含了之后出现显著分化的基因。很明显,人类的基因组是继承和发展的产物,它留下了各种不同进化程度生物物种基因的烙印。这很类似于巴兰金(1988)的一种通俗说法:“比较古老的、原始的、粗糙的机构用各种新的部件进行补充,而在某种程度上又存在于后来的系统之内(例如我们保存着我们类人猿祖先或甚至单细胞祖先的很多品质)”。 有意思的是,一些基因的继承似乎也充满了很大的随机性。譬如,人类蛋白组中有223种蛋白与细菌的蛋白相似,而未能在酵母、线虫、果蝇、拟南芥以及任何其它(非无脊椎)真核生物中找到同源物。进一步的分析表明,人类基因组中至少有113个基因广泛分布于细菌,但在真核生物中仅出现于脊椎动物。很可能编码这些蛋白的基因曾经在早期的原核和真核生物中都出现过,但后来在酵母、线虫、果蝇、拟南芥以及可能还有其它非无脊椎真核生物种系中丢失了。或许还有一种可能的解释是这些基因通过细菌的水平转移进入了脊椎动物(或前脊椎动物)的种系(International Human Genome SequencingConsortium 2001)。
图4 人类同源蛋白的分布(引自International Human Genome SequencingConsortium 2001)
图5 真核生物蛋白组的功能分类(引自International Human Genome SequencingConsortium 2001)
生命界基因的这种传承性也能从动物胚胎发育模式中找到有力证据。早在18世纪,解剖学家就注意到相关种类的动物胚胎之间比它们的成体更为相似,譬如,人类胚胎的早期形态不仅与其它哺乳动物(狗、牛、鼠)胚胎的早期相似,而且在早期阶段甚至与爬行动物、两栖动物和鱼类的胚胎相似;成千上万种动物的胚胎结构有它们祖先的痕迹,但这些相同的结构在成体的生命形态中都没有。后来实验胚胎学家发现,这些具有祖先性状的胚胎结构起着胚胎“组织者”的作用,承接下一个阶段的发育,例如,如果切除一只两栖动物胚胎的前肾管,就不会发育出中肾,同样如果切除原肠顶端的条纹中线,就会阻止脊索和神经系统的发育,因此,“无用的”前肾和条纹中线之所以重演,是因为它们是后期结构发育的胚胎组织者(迈尔2008)。
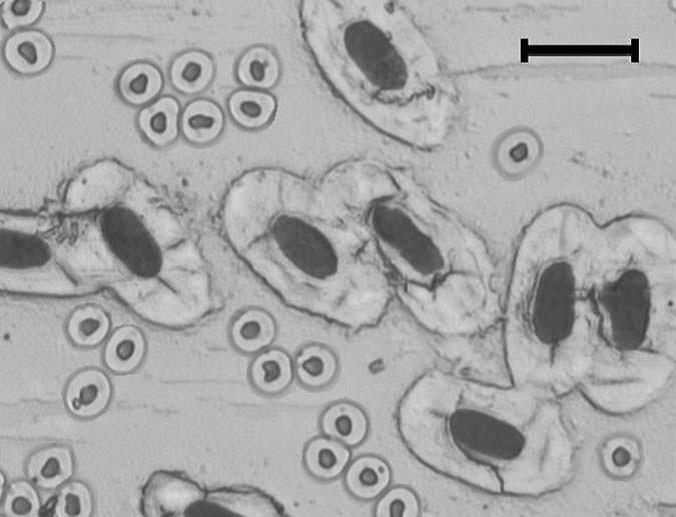
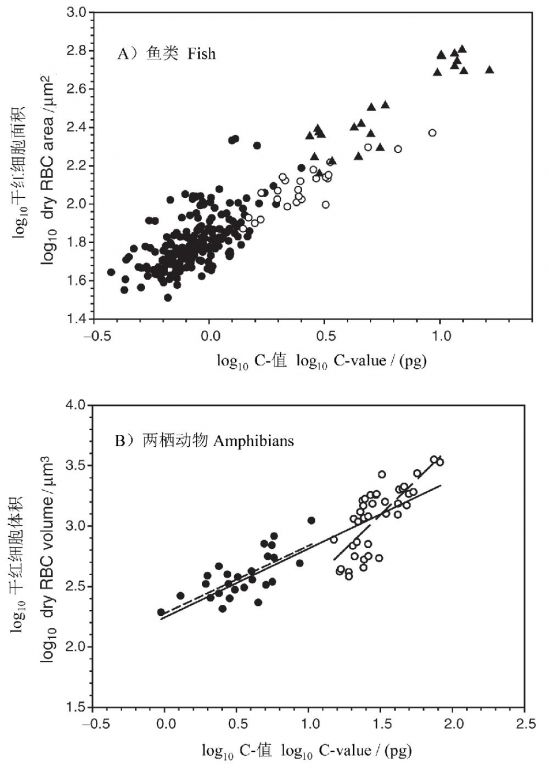
二、基因组大小—折射物种的生理生态对策 1. 大的基因组—需要大的细胞来装填 早在一个多世纪以前,人们就注意到细胞核体积和细胞体积之间明显的正相关关系(Gulliver 1875)。基因组大小和细胞体积大小的关系可用一个极端的例子—图6来说明:这是同时显示的两种鱼的红细胞,肺鱼的核在物理上往暹罗斗鱼的细胞中装都没法装进去!20世纪50年代以来,人们认识到细胞体积是基因组变异的最普遍机制(Gregory 2005)。 在动物中,以脊椎动物的红细胞为材料的研究最多。Hardie and Hebert(2003)收集了230多种硬骨和软骨鱼类的资料,发现基因组大小与干红细胞面积之间存在很好的关系(图7A)。同样,根据Olmo and Morescalchi(1975, 1978)的数据,两栖类的基因组大小与干红细胞体积之间也存在显著的正相关关系(图7B)。类似的关系也存在于爬行动物、鸟类和哺乳动物(Gregory 2005)。
图6 经富尔根染色的暹罗斗鱼(Betta splendens, 2C= 1.3 pg)和澳大利亚肺鱼(Neoceratodusforsteri, 2C≈105 pg)的红细胞显微照片,后者的基因组要大约100倍。照片倍数 x 40,刻度=20 μm(引自Gregory 2001)
图7 脊椎动物红细胞大小与基因组大小之间的关系。(A)鱼类单倍体核DNA含量与干红细胞面积之间的关系:单倍体(●)和多倍体(○)辐鳍亚纲鱼类和软骨鱼类(▲)。(B)两栖动物干红细胞体积与基因组大小之间的关系:蛙(●)和蝾螈(○)(引自Gregory 2005)
2. 大的基因组—更费时间来完成细胞分裂 长期以来,人们就认识到细胞核体积、细胞体积以及细胞分裂周期之间存在密切的关系(Van’t Hof and Sparrow 1963)。一般来说,植物细胞基因组越大,其有丝分裂的周期就越长(图8)。类似的关系也见于被子植物的减数分裂期(Bennett, 1977)。一般会认为,DNA含量越高,合成需要更多的时间,因此细胞分裂周期也会延长。但是,也有报道发现,同一种植物的减数分裂期却随倍性(ploidy)的增加而下降(Bennett and Smith 1972)。
图8 在23°C生长的六种被子植物根尖细胞的DNA含量与最短有丝分裂周期之间的关系,数据源自Van’t Hof 和Sparrow(1963)(引自Gregory 2005)
3.大的基因组—更费时间来完成生命周期 生物个体的发育起决于细胞的分裂与生长,而这必定受到基因组大小的影响。一方面,基因组大小与细胞大小呈正相关,另一方面它又与分裂速度呈负相关关系(Gregory 2005)。那基因组大小对生物个体的发育到底有怎样的影响? 一般来说,大的基因组限制植物的发育速率。Bennett(1972)提出DNA含量可能限制植物最小世代时间(minimum generation time, MGT)(从萌发到最早的成熟种子的产生)的观点,为此,他收集了271个具有不同生活型以及不同MGT的被子植物的资料,比较了各种植物核DNA含量的平均值与范围。他将植物分为4种类型:1)短生植物(ephemerals):能在非常短的时期完成生活史(数周或更短),2)一年生植物(annuals):在52周内完成生活史,3)兼性多年生植物(facultative perennials):能潜在地在萌发52周内产生出可繁殖的种子,4)专性多年生植物(obligateperennials):需要52周以上产生成熟的种子。 核DNA含量(单个染色体):短生植物(1.5 pg) 收藏 IP: 218.249.39.*| 热度| |




【本文地址】
今日新闻 |
推荐新闻 |