不同淡水鱼类游泳速度的初步研究 |
您所在的位置:网站首页 › 鱼类身体呈什么形状的 › 不同淡水鱼类游泳速度的初步研究 |
不同淡水鱼类游泳速度的初步研究
|
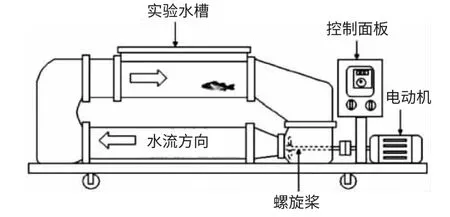
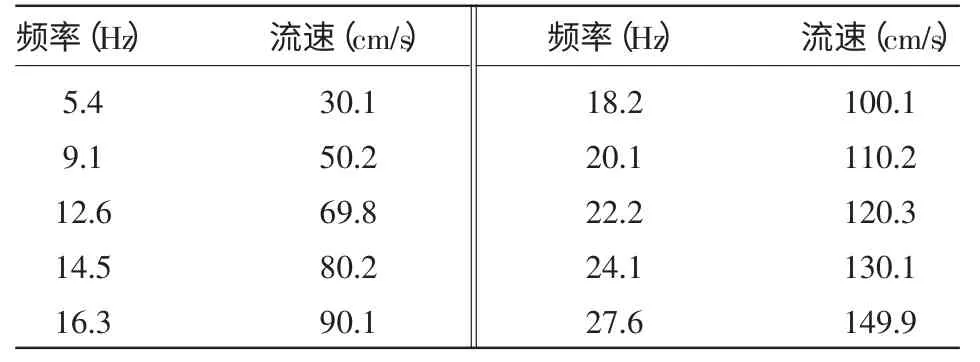
乔云贵,黄洪亮,黄妙芬,徐国栋,殷雷明,陈 帅 (1.大连海洋大学海洋科技与环境学院,辽宁 大连 116023;2.中国水产科学研究院东海水产研究所,农业部海洋与河口渔业资源及生态重点开放实验室,上海 200090) 关于鱼类游泳能力的研究,可追溯到17 世纪,Bainbridge[1]和Wardle[2]等学者分别利用小型循环管道、水槽、池塘、鱼道、鱼梯、排水沟和灌溉等条件做试验,取得了鱼类游泳速度的大量资料。随着科技的发展,He[3]利用水下摄影机观测大西洋鳕鱼(Gadusmorhua)的游泳能力;Kim 等[4]结合模糊数学理论,建立了沿岸底层鱼类的运动模型。而国内研究鱼类游泳行为比较少,井爱国等[5]对花鲈(Lateolabrax maculatus)和许氏平鲉(Sebastes schlegeli)的游泳能力进行了研究;史航等[6]对许氏平鲉、大泷六线鱼(Hexagrammos otakii)临界游速与爆发游速及其生理指标进行了研究。随着技术的发展,人们对鱼类游泳能力研究层次不断深入,鱼类行为这门学科也会日趋完善[7]。 试验研究采用垂直循环水槽分别测量了鲫(Carassius auratus)、鲈鱼(Lateolabrax japonicus)、鳜 鱼(siniperca chuatsi)、罗 非 鱼(Oreochromis niloticus)的临界游速和爆发游速,比较几种试验鱼的游泳能力及其影响因素,以期为渔具开发、鱼道设计和水利工程中过鱼、拦鱼和诱鱼设施制定提供参考数据。 1 材料与方法1.1 材 料1.1.1 试验鱼 将购买的鲫、鲈鱼、鳜鱼、罗非鱼活鱼运至东海水产研究所行为与生理实验室。试验前在曝气自来水的暂养水池(直径为150 cm)中适应2~3 d,24 h 连续供氧,溶解氧6.0~7.5 mg/L,每天换水一次。 1.1.2 试验装置 试验采用新型渔用循环水槽(中国船舶重工集团702 所)(见图1),试验仪器外形尺寸为800 cm×100 cm×285 cm,电机功率15 kW,观测部分为190 cm×50 cm×50 cm。使用流速仪测量流速,得出循环水槽中心水流速度与螺旋桨转动频率之间对应关系见表1。在观测水槽底部标上刻度以测定试验鱼的游泳距离,在观察槽正上方180 cm 和水平方向190 cm 各安装一个摄像头(COLOR CCD CAMERA),视频数据使用灯光及图像系统的硬盘录像机(HIK/DS-8004HF)记录保存。
图1 实验装置示意图
表1 循环水槽观察槽中心的水流速度与叶轮转动频率之间的对应关系 1.2 测定方法1.2.1 临界游速测定 临界游速是描述变速游泳能力所广泛采用的评价指标,试验采用增速游泳方式进行测试,即试验鱼在一定时间步长和流速增长规律下所能达到的最大游泳速度[8]。 暂养结束后,随机选取健康试验鱼1 尾,放入水槽中,在静水适应30 min 后开始试验,记录开始时间。试验鱼在初始流速10 cm/s 游动,若试验鱼可持续游泳20 min,则将水的流速升高10 cm/s,如此反复,直至试验鱼疲劳停止游动,记录试验结束时间。试验结束后将试验鱼从观察水槽中取出,用电子天平称重,测量体长后放入实验室网箱。每种试验鱼9 尾,逐尾计算临界游速(Ucrit),公式如下[9]:
其中,Ucrit为临界游速(cm/s);Up为试验鱼能够持续游泳20 min 的最高流速(cm/s);Ut为流速增幅(10 cm/s);tf为在最后流速段内持续游泳时间(min);ti为时间步长(20 min)。
1.2.2 爆发游速测定 爆发游速是鱼类最快的游速,一般持续时间少于20 s,鱼类通常在捕食、逃避敌害、受到惊吓以及在强水流中游泳时使用爆发游速。 随机选取1 尾试验鱼,静水适应30 min 后,在流动水域中,采用针刺背部或被水流冲至观测槽右侧试验鱼身体撞击栅格的方法,测量试验鱼在受刺激情况下逆流快速前进的游速,直至试验鱼疲劳停止游动。每种试验鱼10 尾,试验录像使用Noldus行为分析软件Ethovision XT 分析计算试验鱼顶流游泳的距离和时间。这时的爆发游速应为水槽流速加试验鱼顶流游泳的平均速度,即[10]:
Uburst爆发游速,Uw为水流速度,S 为试验鱼所游的距离,Ts为试验鱼游 段距离时所需要的时间。爆发游速可以用绝对游速(cm/s)或相对游速(BL/s)表示,即:
试验数据用Excel 2007 进行常规计算后,用SPSS17.0 软件进行多重比较(LSD),分析比较不同体长鲫、鲈鱼、鳜鱼和罗非鱼的临界游泳和爆发游速及之间的差异。统计值用平均数±均值(Mean±SE)表示,差异显著水平为P 2 结果与分析根据鱼类游泳速度公式,经分析和计算得出鲫、鲈鱼、鳜鱼和罗非鱼的临界游速和爆发游速(见表2)。从表2 中可以看出,水温在20.50±1.00℃,体长为16.88±1.99cm 的鲫相对临界游速为7.69±0.59 BL/s,相对爆发游速为8.64±0.60 BL/s;水温在13.02±1.00℃,体长为31.85±2.89 cm 的鲈鱼相对临界游速为2.10±0.14 BL/s,相对爆发游速为2.37±0.11BL/s;水温在18.52±1.00℃,体长为29.46±1.79cm 的鳜鱼相对临界游速为2.50±0.19 BL/s,相爆发游速为4.38±0.24 BL/s;水温在15.57±1.00℃,体长为22.56±2.26cm 的罗非鱼相对临界游速为5.33±0.29 BL/s,相对爆发游速为6.00±0.40 BL/s。
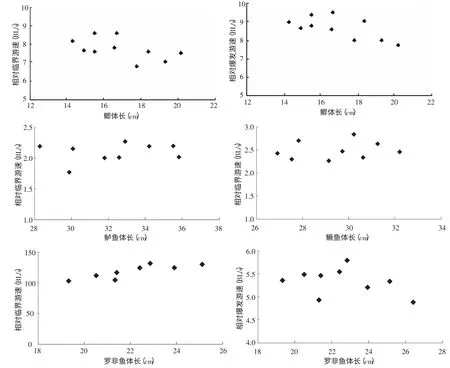
表2 鲫、鲈鱼、鳜鱼和罗非鱼的临界游速与爆发游速 经统计检验得出4 种试验鱼的相对临界游速存在显著性差异(P 从图2 可见,鲫相对临界游泳速度随体长的增长呈递减趋势,鲈鱼和鳜鱼的相对临界游速与体长的关系不明显,罗非鱼的临界游速是随着体长的增大而变大的,而相对临界游速则是相反。
图2 几种鱼的相对临界游速和相对爆发游速 3 讨 论试验鱼在水槽里适应时,常静止在水槽底部或四周,增加水流后,试验鱼移动身体与水流平行,鱼头朝水流方向,不断摆动尾柄、尾鳍和胸鳍,说明这4 种试验鱼都具有趋流性。试验过程中鳜鱼、鲈鱼在水槽中不停的游动,鲫和罗非鱼常常在观察槽的入水栅格处停留。水流较大时,试验鱼上下移动,来回穿越中线的次数明显增加,尾鳍多次撞到栅格上,试验鱼受刺激会快速向前游动,随着撞击次数的增加,顶流前进的距离变短,直至疲劳被水流冲至栅格上。 分析认为试验鱼游泳速度与其体型、体长和生活习性有关。鲫身体呈流线型即纺锤型,尾柄较宽,尾鳍深叉形正尾,这类鱼泳速较高;罗非鱼体短背高,形似鲫鱼尾鳍呈侧弧形,不分叉;鳜鱼的尾鳍圆形;鲈鱼虽然也是纺锤型的,但尾柄长,尾鳍叉形较窄游速较差。试验测定的游速较高,这与游泳能力较强的鱼类具有紧绷肌肉质的身体呈典型的纺锤型及细的尾柄、尾鳍呈镰刀状或新月状等是相符的[11]。 由试验结果可知,4 种淡水鱼类的临界游速和爆发游速是随着体长的增长而增加的:这可能一方面是因为鱼在水中游泳时,其单位体重所需要的推进功率与它的个体大小成反比[12],即随着试验鱼体长增加,大鱼所消耗的运动能量比小鱼的要少;另一方面鱼类生长过程中,肌肉随体长的增加所占总体重的百分比是按比例增加的,自身肌肉收缩所产生的能量也相应增加,所以相同体长的试验鱼游速也有差异。鲫和罗非鱼相对游速则是随着体长的增大而减少的,这与很多学者的研究结果一致[13-14],而鲈鱼和鳜鱼的相对游速随体长变化不明显,可能受到其他因素的影响,有待进一步验证。 鲫是以食植物为主的杂食性鱼类,多在静水或江河缓流中活动,营底栖生活[15];罗非鱼也是以食植物为主的杂食性鱼类,大部分是有机碎屑及其他植物性饲料(如水草类等)[16]。对这种较活跃类型的鱼类,更高的临界游泳能力对其维持日常游泳活动、寻觅食物显得尤为重要,因此,在其整个生活史过程中,较强的有氧运动能力有利于觅食和迁移[17],又因罗非鱼多在池塘网箱中养殖不需要更高的有氧运动能力来捕获食物,游泳速度比野生的河鲫鱼略低。鳜鱼是一种凶猛的肉食性鱼类,伏击取食[18],游泳速度较低,对游至附近的饵料鱼窥视一段时间突发攻击捕食,在网箱养殖中提供较低的水流速度、适口充足的饵料有利于鳜鱼的生长,提高产量。鲈鱼是以摄食底栖小杂鱼为主的肉食性鱼类,活动范围小,喜在静水环境中生活[19],在其生活的水体生态系统中处于较高的营养等级,被捕食压力小,爆发游泳能力对其索饵活动有着更重要的生态学意义。 影响鱼类游泳能力的因素,除了自身因素外还包括温度、光照、溶氧量[20]等。因试验持续的时间较长,受实验室气温影响,循环水槽中的水温在15~25℃波动,不同种类试验鱼一般都在在适温范围内活动,水温越高,新陈代谢越强,肌肉收缩时间截短,故游速也越高,这可能是导致不同种类试验鱼游速差异性显著的外界因素之一。 [1]Bainbridge R.The speed of swimming fish as related to size and to the frequency and amplitude of the tail beat[J].Exp.Biol.,1958, 35:109-133. [2]Wardle C S.Limit of fish swimming speed[J].Nature, 1969, 221:274. [3]He P.Swimming endurance of the Atlantic cod(Gadua morhua L.)at low temperatures[J].Fish Res, 1991, 12(1): 65-73. [4]Kim Y H.Numerical modeling of chaotic behavior for small-scale movements of demersal fishes in coastal water[J].Fisheries Science, 2003, 69: 535-546 . [5]井爱国,张秀梅,李文涛.花鲈、许氏平鲉游泳能力的初步实验研究[J].中国海洋大学学报,2005,35(6):973-976. [6]史 航,陈 勇,赵子仪,等.许氏平鲉、大泷六线鱼临界游速与爆发游速机器生理指标的研究[J].大连海洋学报,2010,25(5):407-412. [7]刘明镜,耿相昌,霍 静,等.鱼类无氧运动及其适应[J].安徽农业科学,2010,38(12):6240-6243. [8]Bellwood D R, Fisher R.Relative swimming speeds in reef fish larvae[J].Mar Ecol Prog Ser, 2001, 211: 299-303. [9]Brett J R.The respiratory metabolism and swimming performance of young sockeye salmon[J].Fish.Res.Bd Can., 1964, 21: 1183-1226. [10]官之梅,刘文郁,陈佩熏.几种淡水经济鱼类游泳能力的研究[C].鱼类学论文集,科学出版社,1981,(1):133-137. [11]何大仁,蔡厚才.鱼类行为学[M].厦门:厦门大学出版社,1988.1-3,12-60. [12]A H 夏皮罗(谈镐生译).形与流-漫谈阻力流体动力学[M].北京科学出版社.1979. [13]Hartwell S I, Otto R G.Critical swimming capacity of the Atlantic silverside(Menidia menidia L)[J].Estuar Coast, 1991, 14(2): 218-221. [14]Petersen R H.Influence of fenitrothion on swimming velocities of brook trout(Salvelinus fontinalis)[J].J Fish Res Bd Can, 1974,31(9):1757-1762. [15]肖 平,陈道印.我国鲫鲤杂交的研究进展及展望[J].江西农业学报,2002,14(3):56-60. [16]李爱景,李学军.尼罗罗非鱼在淡、海水中Na+-K+-ATPase 活性变化[J].河南师范大学学报(自然科学版),2005,33(4):102-105. [17]陈锦云,曹振东,谢小军.温度对瓦氏黄颡鱼幼鱼静止代谢的影响[J].西南师范大学学报(自然科学版),2003,28(4):618-621. [18]李有根,徐济民,黄 平,等.池塘主养鳜鱼高产试验[J].江西水产科学,2011,127(3):32-33. [19]徐 成,王可玲,张培军.鲈鱼群体生化遗传学研究[J].海洋与湖沼,2001,32(3):248-254. [20]Dickson K A, Donley J M, Sepulveda C , et al .Effects of temperature on sustained swimming performance and swimming kinematics of the chub mackerel (Scomber japonicus)[J].Exp Biol,2002, 205 : 969-980. |

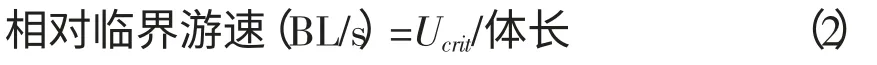

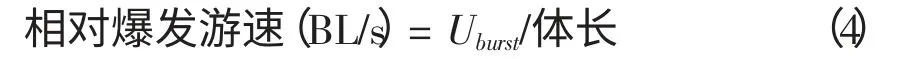
【本文地址】
今日新闻 |
推荐新闻 |