科学网 |
您所在的位置:网站首页 › 染色体决定性别吗 › 科学网 |
科学网
|
Journal Club:性别决定、性别比例和遗传冲突(John H. Werren,1998)
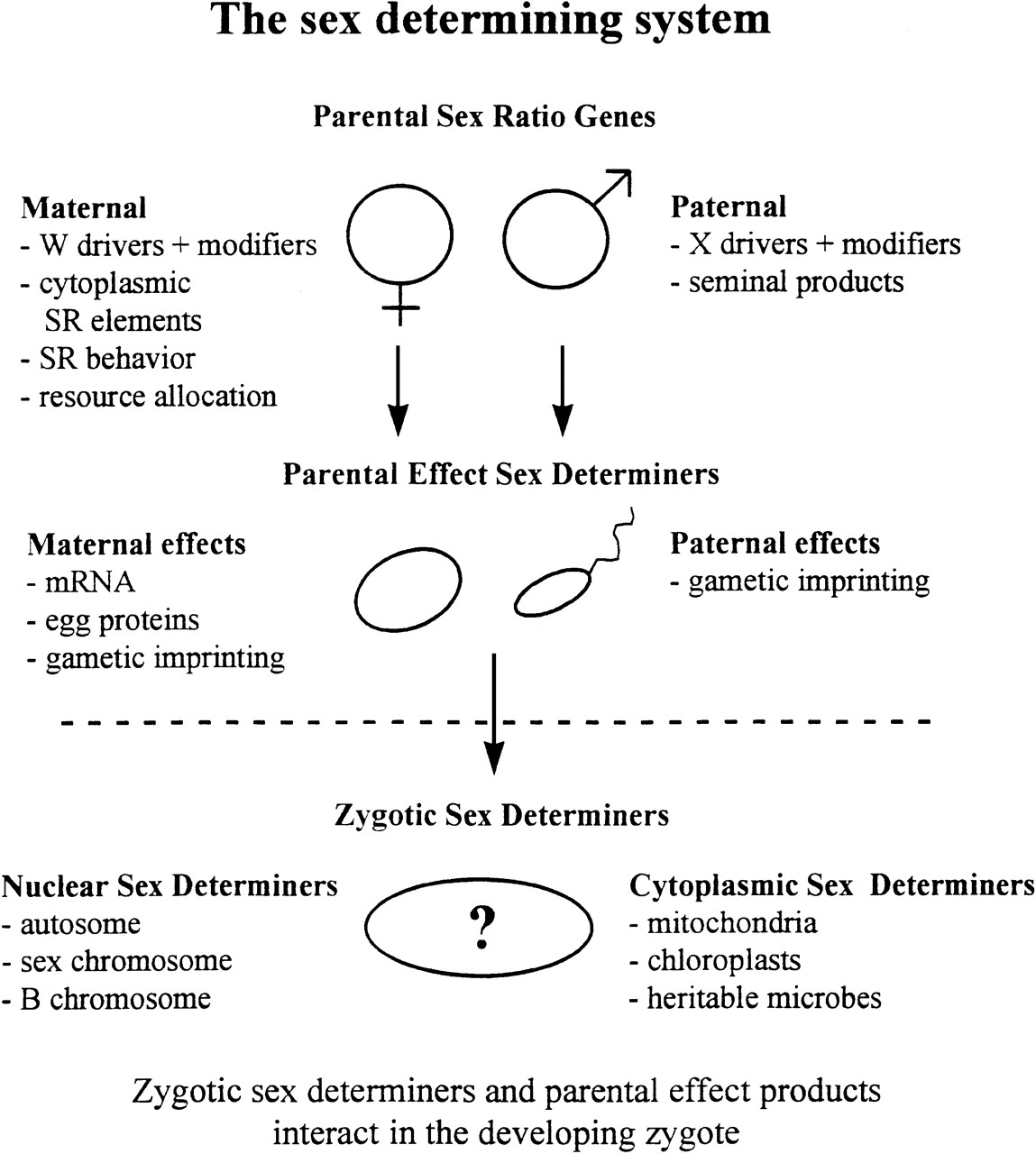
已有 13123 次阅读 2019-1-4 19:07 |个人分类:教学资料|系统分类:科研笔记 性别决定、性别比例和遗传冲突 Annual Review of Ecology and Systematics Vol. 29:233-261 (Volume publication date November 1998) https://doi.org/10.1146/annurev.ecolsys.29.1.233 John H. Werren Biology Department, University of Rochester, Rochester, New York 14627; e-mail: [email protected] Leo W. Beukeboom Institute of Evolutionary and Ecological Sciences, University of Leiden, RA Leiden, NL-2300 The Netherlands; e-mail: [email protected] 摘要 ▪ 摘要 性别决定遗传机制多样性出乎意料,且在进化过程中迅速变化。 我们综述了遗传冲突在多样性和反转背后的驱动力。 当遗传系统的不同组分在相反方向上选择时,就会发生遗传冲突。 基因组之间(包括父系-母系和亲本-合子冲突)或基因组内(细胞质和核基因或性染色体和常染色体之间)都可能发生冲突。 性别决定系统由亲本性别比例基因、亲本效应性别决定因子和合子性别决定因子组成。由于其遗传和表达方式的不同,这些因素会受到不同的选择压力。 遗传冲突理论用于解释几种性别决定机制的进化,包括性染色体驱动、细胞质性比失调和植物细胞质雄性不育。 虽然仍然有限,但越来越多的证据表明遗传冲突在性别决定机制进化中可能很重要。 关键词 基因组冲突;减数分裂驱动;细胞质雄性不育;性别比例失调因子;性染色体 genomic conflict; meiotic drive; cytoplasmic male sterility; sex-ratio distorter; sex chromosome. 看法和概述 植物和动物性别决定机制非常多样化。 一个简短的概述说明了这一点。 在雌雄同体物种中,雄性(小配子)和雌性(大配子)功能都存在于同一个体内,而雌雄异体(或雌雄异株)物种则具有独立性别。 在这些广泛的类别中,性别决定表型和遗传机制进一步多样化。 在雌雄异株物种中,存在各种机制,包括单倍二倍体(单倍体卵产生雄性,二倍体卵产生雌性)、父本基因组丢失(受精后父本染色体丢失的性别决定)、雄性配子异形(具有异形XY性染色体的雄性和具有同形XX性染色体的雌性)、雌性配子异形(ZW雌性和ZZ雄性)、多基因性别决定、环境性别决定和各种其他机制(综述见 17 , 175 )。 性别决定甚至可以在物种内和非常近缘的物种之间明显不同。 例如,新月鱼(platyfish,Xiphophorus maculatus )可以具有雄性配子异形或雌性配子异形( 104 )。 此外,表面看起来相同的机制在基础遗传学上可能存在明显差异。 例如,雄性异配系统可以基于Y染色体上的显性雄性决定子(例如哺乳动物)或X染色体和常染色体上的因子之间的基因平衡(例如Drosophila)。 分子研究表明,参与初级性别决定的基因迅速进化 ( 48 , 111 , 166 , 169 , 170 , 176 )并且一个物种中的性别决定基因可能并不参与近源物种的性别决定( 67 , 100 )。 在这种多样性中存在一个困惑。 虽然人们会认为性别决定这样一个基本的发育方面,在进化中会非常稳定,但事实恰恰相反。 这一观察导致两个重要的进化问题:“为什么性别决定机制如此多样化,性别决定机制如何变化,即如何从一种性别决定机制转变到另一种性别决定机制?”据推测,当某些因素(或多个因素)破坏现有性别决定机制的稳定性时,性别决定系统会发生变化并导致新机制的进化。 因此,重点应放在可能破坏性别决定机制稳定性的因素上,以及性别决定的某些特征是否会使其在进化时间内具有内在的不稳定性。 在这篇综述中,我们考虑了遗传冲突在性别决定系统进化中的作用。 当基因组内的不同遗传元件以在不同方向上“推动”表型被选择时,就会发生遗传冲突。 遗传冲突有两种基本形式。 基因组内冲突涉及生物个体内不同遗传元件之间的选择压力冲突(例如细胞质基因和常染色体基因之间)。 基因组间冲突发生不同个体的遗传元件之间在特定表型上的相互作用。 遗传冲突是性别决定系统的固有特征。 例如,细胞质遗传元件(例如线粒体、细胞质微生物、质体)通常通过卵细胞质而不是通过精子遗传。 因此,对这些元素的选择,将产生强烈的偏雌的性别比例,这增强了它们到后代的传递( 42 , 55 )。 相反,常染色体基因(那些位于非性染色体上的基因)通常被选择产生性别比例平衡( 57 )。 结果,选择细胞质和常染色体基因,会推动性别决定向不同方向进化。 有大量证据表明常染色体和细胞质基因之间的冲突很普遍( 86 , 170 )。 在性染色体和常染色体基因之间以及亲代和后代表达的基因之间也可能发生性别决定的遗传冲突。这些相互冲突的选择性成分之间的共同进化相互作用,可能为性别决定的进化变化提供“发动机”。 我们讨论了性别决定进化的各种模型,重点关注了遗传冲突的潜在作用。 我们认为遗传冲突最可能是性别决定机制多样性的一般解释。 然而,尽管其作用于性别决定的证据越来越多,但尚未确定遗传冲突导致性别决定进化过渡的明确例子。 鉴于此,讨论了未来研究的可能方向。 读者还可以参考有关性别决定机制多样性的综述( 17 , 175 )、性别比例进化( 3 , 31 , 171 )、异形性染色体进化( 27 , 142 )以及果蝇的( 35 , 137 , 156 )、线虫( 36 , 80 )、哺乳动物( 64 , 82 , 100 )和植物( 66 )体细胞和种系性别决定。 历史简述 遗传冲突(Genetic Conflict)遗传冲突的概念与进化生物学中两个密切相关的发展密切相关- 选择对个体遗传因素而不仅仅对个体生物(选择水平)进行操作;观察到某些遗传因素可能是自私或寄生的观点(例如,尽管它们对发生它们的生物体有害,它们仍然具有传递优势)。 关于现在称为基因组内冲突的首批论文包括 Lewis( 110 )的 理论 研究,考虑了植物细胞质雄性不育(CMS)基因的命运,而Howard( 81 )研究了导致动物全雌的细胞质因子。 两者均表明,产生偏雌的细胞质因子可以在种群传递,尽管它们可能会导致极度偏雌的性别比例和种群灭绝。 因此,基因组内冲突的概念从一开始就与性别决定有关。 然而,这些模型对当时自然选择观点的影响尚未得到广泛认可。 植物学家Östergen( 135 )是最早认识到选择可能在基因组的不同部分以不同方向操作的人之一。 在他对B染色体[冗余染色体出现在多种物种中( 102 )]的研究中,他意识到这些遗传元素是寄生的,相对于宿主基因组的其他部分获得了传递优势。 虽然长期受到反对( 127 ),B染色体是自私元素的观点现在已被广泛接受( 7 , 102 , 153 , 173 )。 减数分裂驱动染色体的发现(染色体传递到超过50%的配子)( 150 )也刺激了基因作为选择水平的考量。 通过驱动基因和未连锁抑制因子之间相互冲突的选择压力,可以理解这类系统的进化( 56 , 73 , 94 , 115 , 180 )。 Dawkins( 47 )在推广基因水平上选择操作的概念方面发挥了重要作用。 Cosmides&Tooby( 42 )引入了基因组内部冲突这一术语,并发表了一篇关于基因组内冲突在进化过程中可能发挥的作用的综合论文,包括细胞质遗传、异配生殖进化、雌雄同体向雌雄异体的转变、性别和性别决定的进化。 一些研究探讨了遗传冲突在进化中的作用( 1 , 11 , 55 , 79 )。 通过Doolittle&Sapienza( 50 )和Orgel&Crick( 133 ) 同时发表的文章, 以及自私的非孟德尔元素如转座子、B染色体、细胞质性别比例失调因子等的发现和积累,DNA可能是自私或寄生的想法开始受到关注 。 Werren等(173 )正式定义了自私的遗传因素,并综述了现有的证据。 遗传冲突的概念现在进化生物学中被广泛接受(例如 89 , 118 ; 综述 84 , 91 , 143 )。 最近的理论和实证研究都集中在细胞质和常染色体性别比例因素之间的遗传冲突上( 43 , 59 , 76 , 145 , 148 , 160 , 170 )、性染色体驱动因子与驱动抑制因子之间的冲突( 71 , 72 , 178 )、遗传冲突在性别进化中的潜在重要性( 78 , 89 )、父系与母系基因组之间的冲突与后代资源的分配( 70 )。 虽然其重要性的证据越来越多,但许多现象中遗传冲突在进化中的作用仍有待确立。 性别决定(Sex Determination) 性别决定研究的一个重要早期发展是性染色体的发现( 77 )和异配性别决定理论的发展( 120 )。 随后的研究集中在广泛的生物中性别决定的基本机制(综述于 175 )并发现了相当大的多样性。 性别决定的详细遗传研究仅限于少数生物,最值得注意的是有雄性配子异形的Drosophila melanogaster。 在基因平衡系统中,性别取决于X染色体上雌性决定因子与常染色体上雄性决定因子之间的平衡。 Bridges( 12 ) 在早期遗传实验中发现了这个系统 。他改变了果蝇属物种X染色体的数量,并认为果蝇属性别由X染色体和常染色体之间的比例决定。 在显性Y系统中(例如在一些哺乳动物中),Y染色体上存在显性雄性决定因子。 Bull( 17 )在一篇关于性别决定机制进化的重要论文中,考虑了性别决定系统之间的转换,以及其他 因素之间遗传冲突的可能作用。 性染色体和配子异形的进化也被广泛考虑( 18 , 20 , 26 , 27 , 142 )。 目前,仅有少数生物中详细了解性别决定分子调控,包括家鼠( Mus ),果蝇( D. melanogaster )和线虫( Caenorhabditis elegans )(综述 80 , 156 )。 这些系统可作为与其他系统进行比较的基础。 然而,由于它们的系统发育距离,很难推断导致这些物种之间差异的进化变化。 有关性别比例进化的理论和实证文献非常广泛( 31 , 37 , 63 , 73 , 171 , 179 ),但这些研究大多集中在如何在各种情况下,选择作用于母本操纵后代的性别比例。 很少考虑在亲本中作用的性别比例基因和在受精卵中作用的性别决定基因之间的协同进化相互作用(但请参阅 19 )。 概念框架 性别决定系统(The Sex Determining System) 正如Bull( 17 ) 所指出的, 性别比例选择是塑造性别决定系统进化的潜在驱动力。 性别比例选择涉及通过雄性功能(精子或花粉)与雌性功能(卵或胚珠)遗传因子的传递成功。 当一个特定的遗传因素通过一个性功能比另一个具有更高的传递时,选择将有利于该元素的变体,偏向传递性别的性别比例(或性别决定)。 为了理解性别决定的进化,有必要考虑选择如何作用于整个性别决定系统的每个组成部分。 该系统不仅包括在个体内起作用以确定其性别的基因,还包括在亲本中作用的影响性别比例或性别决定的基因( 图1 )。 性别决定系统的组成部分可以根据其继承方式进一步分类。 遗传因素的遗传方式对性别比例选择如何对其起作用具有重要影响。
传统上,性别决定的遗传研究主要集中在发育受精卵中影响其性别的基因。 然而,性别决定的进化受到作用于三大类基因的选择影响- ( a )性别比例基因,它们是在亲本体内起作用以影响其后代性别比例的基因,( b )性别决定基因,它是在发育受精卵中起作用以影响其性别的基因,以及( c )亲本效应性决定因子,这是在亲本中表达的基因(即它们依赖于亲本基因型)但在发育受精卵中起作用以影响它们性别。 后一类母本效应性别决定基因的例子如 D. melanogaster ( 34 , 156 )、家蝇( Musca domestica )( 51 , 93 ),在表现出全雄性或全雌性家庭的单一性产生的物种中(例如 Chrysomya , 167)、以及在发育早期显示父系基因组丢失的介壳虫( 17 , 130 )。 下面简要讨论这三类性别决定基因,并给出了实例 表1 。 亲本性别比例基因(PARENTAL SEX-RATIO GENES) 很多物种广泛存在亲本对性别比例的影响。 一类亲本性别比例基因驱动性染色体减数分裂。 性染色体驱动是亲本表型,其改变具有X和Y(或Z和W)基因的配子比例,但不直接影响合子性别决定机制。 X染色体驱动已发现于具有雄性配子异形的多种物种中,包括果蝇、蚊子和旅鼠(见下文)。 亲本对性别比例的影响在单倍二倍体昆虫中很常见,其中雌性通过改变卵子受精的可能性来控制子代间的性别比例( 63 )。 未受精的卵发育成雄性,受精卵发育成雌性。 在一些物种中已经记录了受精比例的遗传变异( 135 ),并在许多其他 物种 中有所推测( 63 )。 亲本对性别比例选择的另一个影响机制是对雄性和雌性后代的资源差异分配。 通过为一个性别的后代分配更多资源,亲本表型可以改变对合子性别决定因子的选择。 在具有环境性别决定的物种中,亲本可通过选择产卵位点影响子代间的性别,如水龟( 146 )和西部彩龟( 96 )所示。 反过来,这将影响选择如何操作受精卵中表达的环境性决定基因。 一些鸟类[例如塞舌尔莺(108 )]根据可用资源改变后代之间的性别比例。 这是由于减数分裂过程中,Z或W染色体的优先分离(亲本性别比例效应)或合子性别决定的母本修饰(见下文)。 亲本效应的性别决定因子(PARENTAL-EFFECT SEX DETERMINERS) 在功能上,亲本效应性别决定基因类似于合子性别决定因子,因为它们的产物在发育受精卵中发挥作用。 然而,它们受到与性别比例基因相同的选择压力, 因为它们在亲本中表达并且取决于亲本基因型。 存在母本和父本效应性别决定因子( 表1 )。 大多数母本效应是发育卵子中母本产物(例如mRNA或蛋白质)。 母本效应在早期发育中通常是重要的。在大多数生物中,合子基因型在早期有丝分裂期间不表达,因此早期发育依赖于置于卵中的产物。 因此,由母亲置于卵子中的基因产物可能对发育中的受精卵中的性别决定产生重大影响。 性别决定的分子遗传学研究揭示了几种有趣的母本效应。 在 D. melanogaster ,daughterless( da )是一种母本效应核基因,可产生参与性别决定的转录因子( 34 , 156 )。 M. domestica ( 51 , 93 )和 Chrysomya rufifacies ( 167 )中已经检测到类似的母本对合子性别决定的影响。 Nur( 128 )模拟了母本对性别决定的控制。 父本效应性别决定因子的一个例子似乎是父本性别比例染色体(psr),一种发生在寄生蜂中 Nasonia vitripennis ( 131 )冗余(B)染色体。 通常,这些小蜂通过卵受精(二倍体雌性后代)或抑制卵受精(单倍体雄性后代)来控制其后代之间的性别。通过携带psr的精子对卵子进行受精后,父本染色体(psr除外)在第一次有丝分裂中不能正确凝聚并最终丢失。 因此,受精卵单倍并且发育成雄性。 间接证据表明:尽管其表达发生在受精卵中,psr在精子发生过程中起作用以改变发育精子( 10 )。 尽管目前很少有父本效应的性别决定因子,但它们可能比我们认识到的更为常见。 一种机制可能是性别决定基因的父系印记,从而影响它们在发育受精卵中的表达( 8 , 125 )。 合子性别决定因子(ZYGOTIC SEX DETERMINERS) 性别决定的经典研究认为在受精卵中起作用的基因确定其性别。 合子性别决定因子的例子包括: 老鼠和人类中的SrY ( 64 )、 D. melanogaster 中的Sex lethal ( 156 ),和C. elegans中的 XOL 和 SDC 基因 ( 80 )。 同时 D. melanogaster 和 C. elegans,主要的性别决定信号是X:A比率。X染色体上存在多个X分子元素,涉及几个基因的调控级联决定了躯体性别( 80 )。 X:A系统的进化似乎与剂量补偿进化有关。 一个尚未解决的进化问题是X:A性别决定是从一个祖先的状态进化而来的,可能涉及一个新生性染色体上的主要性别决定因子。 换句话说,为什么系统从主效应基因进化为X染色体上的多个雌性决定因子和常染色体上的雄性决定因子? Wilkins( 177 )基于这些系统的分子遗传结构提出:C. elegans和 D. melanogaster 性别决定是通过顺序添加遗传开关而进化的,每个都逆转了之前的性别决定。 他进一步提出,这一过程是由频率依赖的性别比例选择驱动的。 该模型与遗传冲突或其他机制引起的性别比例强选择一致(见下文)。小鼠和人类中 存在一种显性的雄性决定因子( SrY ),尽管目前尚不清楚 SRY 是否是主要信号或其他信号是否诱发 SRY 睾丸决定级联( 100 )。 性别决定的遗传冲突(Genetic Conflict Over Sex Determination) 当性别决定系统的各个组成部分被选择,以在不同方向上推动合子性别决定或亲本性别比例时,将发生遗传冲突。 鉴于作用于不同遗传模式(细胞质,常染色体和性染色体)和不同表达位点(母本,父本和合子)的基因作用的选择压力不同,遗传冲突是性别决定系统的固有特征。 在这里,我们列出了性别决定和性别比例冲突的一般领域。 细胞质-核冲突(CYTO-NUCLEAR CONFLICT) 细胞质和核基因之间在性别决定和性别比例方面的冲突是显而易见的,并且似乎是普遍存在。 许多细胞质性比例失调因子是通过卵细胞质而不是精子传递微生物(综述于 86 )。 在植物中,已经记录了母系遗传细胞器诱导CMS和细胞质雄性不育(CMS)的常染色体抑制因子之间的细胞质-核冲突(综述于 39 , 148 )。 在没有抑制或其他平衡力的情况下,细胞质性别比例失调因子可能在固化点附近或扩散,可能使种群(和物种)灭绝( 81 , 160 )。 下面将更详细地讨论细胞质-核冲突。 性染色体驱动和B染色体驱动冲突(SEX-CHROMOSOME DRIVE AND B-CHROMOSOME DRIVE CONFLICT) 性染色体驱动只是有利于减数分裂驱动基因位点的选择的一种表现,也发生在常染色体上(综述于 114 )。 然而,由此产生的性别比例失调会产生强烈的性别比例选择。 有大量证据表明X染色体驱动选择Y染色体和常染色体上的抑制因子(见下文)。 在性染色体上重组的物种中,对连锁基因的选择有利于驱动的增强或抑制。这取决于基因的连接紧密程度以及是否维持连锁不平衡( 180 )。 然而,X染色体驱动引起的性别比例失调,可能有利于合子性别决定(或母本效应性别决定因子)的补偿性变化,还尚未被广泛研究。 性染色体驱动也可能导致种群灭绝( 73 , 112 , 113 )。 许多B染色体是寄生性遗传因子。即使它们对宿主施加适应性成本,染色体仍保持在种群内,增强它们在配子(传递驱动)中的传递,( 127 , 129 )。 在许多情况下,Bs通过雄性和雌性(或雌雄同体中的雄性和雌性功能)的传递是不对称的。 在这种情况下,预计选择会导致性别比例和性别决定基因的积累,从而使性别偏向于传递性别。 然而,详细研究具有B染色体偏向传递的少数介壳虫物种,未能显示B染色体对性别决定的影响(U Nur,个人通信)。 性别比例失调B染色体的一个突出例子是先前描述过的 N. vitripennis 的psr 染色体 ( 131 )。 亲本-后代冲突(PARENT-OFFSPRING CONFLICT) Trivers( 163 )最初提出了这样的观点,即亲本和后代可能具有不同的遗传利益,因为它们遗传近缘但并不遗传相同的事实。对亲本- 子女冲突的研究通常涉及分配给后代的资源数量的冲突。 然而,Trivers&Hare( 164 )提出,在社会昆虫中,女王与其后代工虫之间应存在性别比例冲突。 实证研究为这种冲突的存在提供了强有力的支持( 159 )。 鉴于母本效应性别决定基因的证据越来越多,需要更彻底地考虑性别决定冲突的可能性。 有两种情况可能发生这种冲突:( a )当雄性和雌性子代的亲本的适合成本不同时,以及( b )部分近亲繁殖或局部配偶竞争(LMC)。 当亲本生产一个性别成本高于另一个性别时,自然选择将有利于亲本过度生产成本较低的性别( 57)。 然而,对受精卵的选择通常有利于更平衡的性别比例。 当母本的成本是未来的生存和繁殖时,尤其如此。 例如,在马鹿( Cervus elaphus )中,生产雄性对于雌性母本来说比生育雌性更具生殖成本,并且雌性母本经常在雄性出生后的一年内无法繁殖( 38 )。 理论上尚未探索这种相互作用的动态。 根据交配系统,父本效应性别决定者将具有与合子或母系基因更一致的遗传兴趣。 在部分近亲繁殖或局部配偶竞争中,母本效应基因将被选择以产生更多偏雌的性别比例。合子效应性别决定因子 也将被选择偏雌,但由于遗传相关性的不对称,均衡比率应该会较少偏向。 结果将是相互冲突的选择压力。 可能的结果将是母本修饰物和合子修饰物在相反方向上的积累。 同样,理论上还没有探索过相互作用的系统。 亲本性染色体驱动因子与合子性别决定基因之间也存在冲突。 原则上,驱动性染色体导致的性别比例失调会导致性别决定向代表不足的性别补偿性转变( 113 )。 母系-父系冲突(MATERNAL-PATERNAL CONFLICT) 研究兴趣主要集中在母系和父系衍生基因之间的基因组内冲突,而不是资源分配给发育受精卵和基因组雌雄冲突对雌性生殖努力的影响( 70 )。 然而,性别决定的进化有一些有趣的应用。 Brown( 13 )和Bull( 15 岁 , 17 )已经表明,母系基因/父系基因冲突可导致父系基因组丢失和单倍二倍体性别决定的进化。 基本上,母系基因具有“消除”父系基因组的选择性优势。 这种优势(称为Brown的自动频率响应)是由于下一代单倍体雄性中的母本基因组传递相对于二倍体雄性的更高(即由于减数分裂而没有减少)。 只要单倍体雄性适合度大于二倍体雄性的一半,这种优势就会增加。 此外,在具有单倍二倍体和父系基因组丢失性别决定的物种中,基因组母系- 父系冲突明显发生( 71 )。 在单倍二倍体中,选择雄性以增加其配偶产生的受精卵比例(雌性比例)。 但是,目前尚不清楚雄性有哪些机会可以影响雌性的性别比例。 在 父系基因组丢失系统[例如介壳虫( 130 )]中,父系基因被选择以逃避或抑制父系基因组丢失。 一些冗余的染色体已经从父系基因组丢失中进化出逃避机制,例如在粉虱( 127 )和扁虫(Polycelis nigra )( 9 )。 性别决定进化的替代模型 遗传冲突是性别决定系统的固有特征。 然而,除了遗传冲突之外,还提出了许多用于性别决定进化的模型。 我们简要回顾一下目前文献中的一些模型,重点关注那些破坏性别决定系统稳定性并导致性别决定机制进化的因素。 短时适合度和性别协变(TRANSIENT COVARIANCE OF FITNESS AND SEX)(HITCHIKING) Bull( 17 )提出,在强阳性选择下,性别决定等位位点和基因之间的瞬时连锁不平衡,可能通过引起群体性别比例失调来破坏性别决定的稳定性。 这些失调的性别比例会对产生异性的性别决定位点产生反选择。 这种影响可以解释在 M. domestica中 发现的性别决定多样性 ,其中一些性别决定变异似乎与杀虫剂抗性等位基因关联( 106 , 124 , 147 )。 在新月鱼中,几种体色基因与性别决定基因位点紧密关联( 104 )。 积累-消耗(ACCUMULATION-ATTRITION) Graves( 67 )提出了一个“积累- 消耗”模型来解释哺乳动物性别决定进化。 根据该模型,哺乳动物性别决定通过一系列常染色体添加(易位)到Y染色体,随后是这些假常染色体区域退化,实现进化。当 X和Y染色体之间的 交叉被抑制时会导致突变降解 ,而只有雄性性别决定中进化功能的基因才能逃避突变降解。 一系列易位事件可导致Y上的性别决定基因反转。该模型与性拮抗基因可在性染色体上积累的观点(例如,Y连锁基因增强雄性适合度并降低雌性适合度)( 141 , 142 )和雄性生长促进剂会在Y上积累的观点( 87 , 88 )一致。 种群结构与近交(POPULATION STRUCTURE AND INBREEDING) Hamilton( 73 )指出,局部交配(和近亲繁殖)的细分种群选择具有偏雌的性别比例的亲本。 有大量的实证证据表明,区部配偶竞争确实导致了偏雌的性别比例(综述于 3 , 74 )。 然而,在没有亲本性别比例控制的情况下,几乎没有考虑近亲繁殖和当地配偶竞争如何塑造物种合子性别决定机制。 与性别决定进化相关的另外两种种群结构效应是当地资源竞争( 33 )和当地资源增加( 152 )。 因雄性和雌性(或雌雄同体的雄性和雌性功能)不同,每当适合度回报作为该性别投资量的函数(例如由于差异扩散),将选择有偏向的性别比例( 58 , 59 )。 然而,大多数这些影响的模型隐含地假设了亲本性别比例控制。 同样的选择力也应该选择合子性别决定基因偏向,尽管强度会低于亲本性别比例和亲本效应性别决定基因。 这种效应在理论上尚未得到研究。 雄性和雌性可变适合度(VARIABLE FITNESS OF MALES AND FEMALES) 当雄性和雌性适合度受到某些环境因素的不同影响时,预计会对性别比例和性别决定进行兼性调整。例如,Trivers&Willard( 165 )指出:当母本状况发生变化时,这种变化会转化为对 雄性子代和雌性子代 的更大的适合效果;选择将有利于母本状况良好时过度生育雄性子代,而母本状态不好时过度生育雌性子代。可变适合度效应还可以解释顺序雌雄同体的年龄特异性变化和宿主大小对寄生蜂性别的影响(31 )。 可变适合度效果在环境性别决定进化几乎肯定是重要的( 16 , 32)。环境性别决定在某些海洋蠕虫和软体动物观察到的:寄生线虫如mermithids、一些植物(136)、少数鱼、一些蜥蜴、海龟和鳄鱼(综述17,97)。在无脊椎动物中,拥挤或营养不良通常与雄性决定增加有关。性别决定在各种爬行动物中对温度敏感,尽管有利于这种性别决定的选择性因素仍不清楚。 锁定性别决定(Locked-In Sex Determination)?一些性别决定系统可能比其他系统更严格,从而减少或阻止系统的进一步进化。异形性染色体被认为是同形性染色体之间重组抑制后,杂合状态下的染色体突变变性(Y在XY雄性和W在ZW雌性)主要进化而来( 27 , 142)。一旦异形性染色体进化,性别决定的进一步变化可能受到XX雄性,XY雌性和/或任何性别的YY个体的不育或不活的限制(21)。例如,在小鼠和人类中,Y染色体上存在雄性可育因子,限制了XX雄性的潜在适应性(67)。系统发育模式支持性染色体异型性进化增加了性别决定机制保守性(17,133)。 性别决定基因的多效性可以限制性别决定进化。例如,性别决定和剂量补偿之间复杂的相互作用可能会限制异形XX / XY 和ZW / ZZ性别决定系统变化能力。由于剂量补偿和初级性别决定与 D. melanogaster 的X:A平衡系统密切相关,中性性别决定基因中的突变体,性致死(Sxl ),因为剂量补偿中断对于一种性别(因此名称)通常是致命的(149)。在人类中,SrY 和相关性别决定基因(DAX1 ,SF1 )对其它发育过程,如骨骼、神经和肾上腺发育具有多效性效应(105,140)。 反对的观点,即性别决定机制可被锁定。越来越多的证据表明,表面上相似性别决定机制可以有潜在的不同的遗传结构( 41 , 111,126)。例如,鼠类啮齿动物物种的SrY 基因数量不同(111),并且即使在不同地理种群的Mus musculus 之间,SrY的效力也不同,导致在种群内杂交中产生雌雄同体和XY雌性(126)。此外,很明显,即使被认为异形性染色体(例如哺乳动物)保守的种群也会显示出该性状变异。先前认为具有遗传性别决定系统的一些脊椎动物实际上由遗传和环境性别混合决定(44),并且这些机制之间的过渡可以是相对容易的(40,45,101)。 遗传冲突系统(GENETIC CONFLICT SYSTEMS) 性染色体驱动(Sex-Chromosome Drive) 减数分裂驱动染色体以非孟德尔方式遗传,通常以70-100%的配子结束( 150 )。最著名的例子是Drosophila分离失调(46,162)和Mus的t-locus(112)。减数分裂驱动性染色体很容易识别,因为它们对后代性别比例有直接影响。它们见于几个哺乳动物和昆虫类群,包括果蝇、蚊子和蝴蝶(综述92)。大多数例子是驱动X染色体,通常称为性别比例(SR)染色体。驱动Y染色体是罕见的,可能是因为它们更强的驱动能力导致在没有反选择的情况下快速灭绝(73)。 最近的证据( 5 , 94,122)与Frank(60)和Hurst&Pomiankowsky(92)预测相符:驱动性染色体比以前认为更为常见。如果没有反选择,性染色体的减数分裂驱动将很快导致携带者种群灭绝(73)。反选择可以发生在基因、个体和种群层面(见94)。在个体层面,驱动性染色体通常会降低雄性的生育能力(115)。该行为模式的结果,通常涉及携带非驱动性染色体同源基因的配子功能障碍(138)。如果驱动基因与染色体倒位相关,那么雌性也可能降低适应性(见95)。Wilkinson等(178)发现Y驱动的频率随着眼柄增大的种群中相关反应而增加。这表明涉及该雄性角色的基因是Y连锁的。 选择通常有利于常染色体上的等位基因和抑制SR染色体的减数分裂驱动的非驱动性染色体。理论模型表明,性染色体驱动的系统最有可能发展成一个双位点多态性与连锁不平衡( 114 , 180)。预计驱动等位基因显示与增强子等位基因的偶联和抑制等位基因的排斥,这可能通过染色体倒位进一步促进(115)。常染色体抑制因子向驱动的进化并非不可避免,这取决于雄性和雌性驱动染色体的特定适应性效应(180)。Jaenike(94)在没有连锁的情况下引起了频率依赖性选择。许多生物发生SR染色体修饰(5,23,24,114,122,139,155)。例如,Cazemajor等人(25)表明,在Drosophila simulans ,驱动的结果来自几个X连锁基因位点的作用以及在每个主要常染色体上以及Y 染色体上驱动抑制因子的驱动修饰。类似地,在植物Silene alba中,Y染色体上的恢复基因位点平衡了由假定的驱动X引起的性别比例偏差(161)。Hurst(85,90)认为:D. melanogaster 的Stellate 位点是X染色体上的遗留驱动基因,已经被Y染色体上修饰基因沉默。 驱动性染色体清楚地说明了基因组内冲突。然而,性染色体驱动是否选择了合子性别决定机制的补偿性变化?这在自然界没有强有力的证据。所有已知的修饰基因似乎都抵消了母本内驱动作用。相比之下,Lyttle( 113 )构建了驱动Y染色体的实验室群体,其中含有 D. melanogaster 中的分离失调(SD)基因。在大多数种群中,驱动抑制因子进化,但在一个种群中,性别比例失调被性染色体非整倍体(XXY雌性和XYY雄性)的积累所抵消。这个例子表明,一个新的性别决定系统(尽管保持X:A比率)可能会随着驱动性染色体的变化而进化。需要更多这样的实验来探究性染色体驱动的可能进化结果。性染色体驱动是否选择合子性别决定系统的变化可能取决于种群中性别比例失调的严重程度以及相关性状的遗传变异的性质。 动物细胞质性比失调因子(Cytoplasmic Sex-Ratio Distorters in Animals) 细胞质遗传性别比例失调因子在动物中广泛分布(综述 54,83,86)。在大多数情况下,细胞质性比失调是由母系遗传的微生物引起的,这些微生物会改变性别比例并使其偏雌。细胞质性别比例失调因子包括雄性致死、主要性别比例失调因子、雌性化和单性生殖诱导。雄性致死微生物的实例包括: Drosophila willistoni 的螺原虫(spiroplasms) (69)、Nasonia小蜂的 γ-变形杆菌(174)、立克次体、瓢虫的螺原体和和黄杆菌(83,172)以及蚊子的微孢子(2)。遗传雄性的雌性化是由等足动物中的Wolbachia 立克次体(144)和端足动物中的微孢子虫引起的(52)。在一系列膜翅目昆虫中发现了Wolbachia 诱导的孤雌生殖(158 ;在157中综述),并且与其他生物有关。Nasonia中的msr元素引起对雌性的主要性别比例改变(154); 尽管未知导致的因子,但可能是由于线粒体变异。 细胞质性比失调因子和核基因之间的协同进化相互作用可能很复杂。当性别比例失调因子的传递不完整时,亲本性别比例的补偿性变化被选择,可以导致产生单一性别的正反馈- 一些雌性产生全雌性后代(细胞质控制)和一些产生全雄性后代(补偿后核控制)( 17 , 170)。当失调因子的传递接近100%(170)时,不会发生这种效应。对于细胞质性别决定因子也显示出类似的效果(53)。由于性别比例选择,细胞质失调因子的常染色体抑制因子在亲本和受精卵中都是有利的(168)。尽管预计进化出了雄性杀手的抑制因子,理论研究表明至少在随机交配种群中,没有补偿性别比例等位基因选择以应对雄性致死微生物(170)。 Taylor( 160 )研究了合子核性别决定因子(代偿基因)、合子抑制因子和细胞质雌性化因子的协同进化。他发现补偿性核雄性决定因子会增加。然而,在核恢复系统存在的情况下,性别比例通常会进化回1:1,并抑制细胞质因子。如果这个过程在自然界中常见,那么种间杂交可能揭示细胞质性比失调,因为它们从抑制基因型中释放出来。已经提出,杂交致死和不育可产生这样的结果( 60 , 92)。细胞质性别比例失调因子的一个有趣特征是通过关联线粒体搭便车。如果失调的传递不完全或存在恢复基因,则与细胞质失调因子相关的线粒体变体可以在种群中固定。类似的论点适用于植物的细胞质不育(见下文)。可以限制细胞质失调因子扩散的性状包括YY个体的适应性降低(在雄性异配系统中)(160)和针对具有雄性稀缺性的当地种群的群间选择(17)。 尽管有预期,但没有广泛的经验证据证明动物性别决定的细胞核-质冲突。然而,很少有系统被详细研究过。最好的例子发生在 等足动物 Armadillidium vulgare。其群体中可能存在雌性化的Wolbachia 、第二个未知病因学的雌性化因子(f )、雄性化常染色体基因和雌性化因子抑制因子(145 ;见144)。该 f 因子显示复杂的遗传模式,主要是细胞质传递,但也有一些父系传递。 Wolbachia 和 f 之间的明显关联 使Legrand&Juchault( 109 )提出这个想法: f 是一种细菌噬菌体,携带来自 Wolbachia的雌性化元素,偶尔掺入等足动物 基因组。目前还不清楚f 是否实际上是一个显示可变外显率和表达的核基因。一种占优势的雄性化基因已被表征,可以在f存在的情况下恢复雄性;但在雌性化的 Wolbachia (主要是导致功能性的双性)存在的情况下,这种情况只是微弱的。尽管雌性化因子的存在与雄性化的常染色体基因相关,但这些元素的频率存在种群显著差异。 A. vulgare 通常具有雌性异形(ZZ雄性:ZW雌性)。然而,在具有雌性化因子的群体中,由于性别比例的选择,雌性决定性染色体(W)可以从种群中驱动。Juchault&Mocquard(103)提出了一个循环,其中存在不完全传递的Wolbachia 导致W染色体的丢失,导致ZZ雄性和ZZ + WO雌性随后整合 f 因子到常染色体上,产生新W(雌性决定)染色体。这个过程将有效地防止退化(异形)性染色体的进化。还 不清楚的是,核质细胞冲突是否会导致性别决定从雌性配子异形转变为雄性配子异形(即由于常染色体雄性化因子扩散和雌性化因子抑制因子)。尽管理论上还没有探索过各种可能性,但事件的顺序很可能会严重影响这种遗传冲突的结果。Rigaud( 144 )指出,性别决定(雄激素腺体的产生)的生理机制可能使等足类动物特别容易被细胞质元素“劫持”性别决定。 植物细胞质雄性不育(Cytoplasmic Male Sterility in Plants) CMS是由细胞质遗传因子引起的花药或花粉败育。CMS广泛存在(例如玉米、 Petunia、水稻、菜豆和向日葵),并在物种内具有多态性,是具有雌雄同体和雄性不育个体的混合物(称为雌蕊)。Lewis(110)首先指出,当由细胞质而不是核基因引起时,更容易选择雄性不育。只要雄性不育植物比雌雄同株植物产生更有效的胚珠,CMS将受到选择性青睐。例如,当存在胚珠生产的资源分配或(甚至轻微的)雄性不育胚珠的远系繁殖优势时,就会发生这种情况。相反,只有当产生超过两倍的有效胚珠时,才有利于显性核雄性不育基因。其结果是核质冲突,现在有大量证据表明,这样的冲突在植物中发生(43,49)。 这种冲突表现为CMS基因与CMS的核抑制因子之间的复杂相互作用。由于许多显示CMS的植物物种具有重要的经济意义,因此对CMS进行了广泛的分子遗传学分析(综述于 148)。在所有研究过的例子中,CMS基因发生在线粒体内,并且是由遗传重排产生的嵌合体。已显示雄性育性的核恢复体,通过消除CMS序列(在Phaseolus vulgaris菜豆中)和修饰CMS转录本(在玉米中)或转录本丰度(在矮牵牛中)而起作用。 遗传学研究表明CMS基因与核恢复系统在许多系统中具有特异性( 107 )。大多数雌花两性花异株物种具有不止一种CMS细胞型和多种相互作用的核恢复物在种群内分离。例如, Plantago lanceolate(车前草)有三种不同的CMS类型,每种都有一组特定的核恢复基因位点(49)。它们包括从显性到隐性到表面相互作用的恢复者。可能恢复基因的发生抑制了CMS细胞型在许多物种的传递,尽管其它过程如同类群选择也可能涉及其中(59,119)。在某些情况下,CMS细胞型可以在一个物种内固定,但会被恢复等位基因抑制,因此是隐存的。随后可以在种间杂交中检测到这种情况,其中CMS细胞型逃脱其核抑制。与此情景一致,CMS是植物杂种不育的常见来源(110)。 有广泛的理论文献研究CMS和核基因的协同进化动力学(例如 28,29,30,59,65,116,119,151)。其中有趣的问题是,雌性两性异体是否是雌性异株进化的一个过渡阶段,即细胞核-质冲突是否会促进雌性异株的进化。与此观点一致,Maurice等人(116)记录了雌性两性异体和雌雄异体的分类学关联。一种建模研究方法涉及雌花两性花异株种群中核雌性不育等位基因的命运(极端形式的补偿基因)。结果普遍表明,雌雄异株的进化受限,但有可能发生(28,30,116,151)。需要更多的模型来确定是否可以通过雌花两性花异株种群中性别分配到雄性功能的连续变化,而不是通过雌性不育等位基因效应来进化。同受到青睐并认为向雄性功能性分配移位的观点一致,Atlan(4)在百里香(Thymus vulgaris)雌性两性异体种群中观察到这种性别分配偏移。因为表型模型不能捕捉到等位基因的非随机关联(配子相不平衡),而这对于不同基因型的最终命运至关重要。研究这些复杂过程所必须显性遗传模型(例如30)。然而,在这些植物系统中存在最明显和最引人注目的例子,表明导致性别决定等位基因翻转的遗传冲突。 其他系统 在下述情况下,遗传冲突被激发为性别决定系统进化的驱动力。这些系统表明遗传冲突的作用( a )在大多数情况下仍然是假设的,( b )由于某些情况下缺乏信息而无法完全解释,( c )因为它可以帮助解释遗传性别决定系统结构值得考虑,。 旅鼠(LEMMINGS) 旅鼠异常性染色体系统进化已被广泛考虑( 61 , 62,68)。木旅鼠(Myopys schisticolor )有三种类型的个体:XX是正常雌性,XY是正常雄性,X * Y是雌性。变体X染色体[X * (被认为抑制Y的雄性确定效应,使得X * Y个体为雌性)]显示X * Y雌性的驱动,这导致对具有此类型染色体种群中强烈偏雌的性别比例。X * Y雌性通过不分离(YY细胞死亡)具有X * X * 卵母细胞,并产生几乎所有雌性子代。已经从各种旅鼠(Dicrostonyx groenlandicus 和 D. torquatus )描述了这种类似的系统( 62 )。在这些物种中,X * Y雌性也会出现,但大概是通过产生X * 和Y卵,产生雄性子代和雌性子代,。X * 被认为是抑制Y染色体上的雄性决定因子。 一些作者模拟了这些系统的进化动力学和考虑选择会如何导致生殖生物学变动( 6 , 19,22,117)。大多数工作涉及细分种群结构下,近亲繁殖和降低生育率的如何影响驱动X * 染色体及其潜在抑制因子的传递(综合论述见19)。X * Y雌性在多大程度上选择性别决定系统的变化?消除XY雌性最直接的方法之一是X * 的Y连锁抑制基因的进化,但抑制因子Y的入侵在近亲繁殖中受到限制。这与在旅鼠种群中存在抗性Y的经验证据很少的事实一致。结构化种群模型进一步表明,常染色体恢复基因的选择甚至弱于Y连锁抑制。木旅鼠中的实际进化路径似乎涉及X *的进化:雌性化的X * Y雄性随后是分离比例的修饰因子的进化,因此X * Y雌性仅产生X * 卵母细胞,克服了它们的生育能力降低(当Y精子受精时,有一半的卵母细胞死亡)。另一种情形,McVean&Hurst(121)认为目前的情况是对驱动Y染色体的反应,即X * Y雌性通过抑制其雄性决定基因并仅产生X * 卵母细胞来抵消驱动Y的扩散。然而,没有经验证据表明存在驱动Y。总之,旅鼠异常性染色体系的进化可能是从遗传冲突的角度来解释的,但其确切的作用尚不清楚。 菌蚊(SCIARA COPROPHILA) 在菌蚊 Sciara coprophila 中,性别决定与父本基因组丢失相关(14,123)。所有受精卵最初都是XXX,性别由母本因素决定,导致体细胞丢失X染色体。此外,某些染色体(所谓的有限的或L染色体)存在于种系中,但不存在于体细胞中。在精子发生过程中,除了L染色体和母本X(其加倍)之外,所有父本衍生的染色体(即X和所有常染色体)都被消除。因此,雄性仅传递母系衍生的染色体(即两个X和所有常染色体)。除了在早期发育期间被消除的一个父系X,雌性传递所有父系和母系衍生的染色体。 Haig(72)提出了一种基于基因组冲突的进化方案来解释这种不寻常的性别决定机制。他设想了以下步骤:(a )驱动X染色体的起源导致偏雌的性别比例,通过(b )消除一个父本X将XX雌性子代转变为雄性子代来抵消,以及(c )起源来自X染色体且可有可无的L染色体的,有利于偏雄的性别比例,其次是(d )X'染色体的起源,抑制L染色体的效应。冲突的各方是驱动X染色体和L染色体,通过偏向雌性和母本常染色体以及变异(加倍)X抵消其效应,从而获得传递优势。对这种情形的弱测试预测L染色体来自X染色体。 介壳虫 已在遗传冲突的背景下描述了( 13 , 15,71,75)介壳虫(Cocoidea)异常染色体系统的进化( 130 )。使用与Sciara 相似的推理,Haig(71)试图通过一系列进化过渡来解释配子异形中父系基因组丢失的起源。在介壳虫发现了一些这类短暂阶段,许多物种表现出父系基因组中丢失[因为父系染色体是在不同发育阶段的胚系的淘汰,它们不被雄性传递(见14,130的细节和引用)]。Haig的模型包括三个步骤:(a )XO雄性X染色体的减数分裂驱使偏雌性别比例; (b )雄性母系常染色体的连接以利用X-驱动; (c )通过母本表达的常染色体基因将XX雌性子代转变为雄性子代。一个结果,如在Sciara中观察到的那样,母本可能通过控制胚胎发生过程中X染色体的消除来确定其后代的性别。性染色体驱动与常染色体抑制因子之间的冲突被认为是驱动力。Haig的模型说明了遗传冲突如何导致新性别决定机制。尽管在进化上似乎是合理的,但目前还没有经验证据支持表明观察到的系统确实是性别决定基因之间冲突的结果。 鼹鼠(MOLES) 应用遗传冲突理论,McVean&Hurst( 121 )提出了三种进化途径来解释鼹鼠间性体的高频率( Talpa europaea) 和 T. occidentali)( 98 , 99)。雄性是XY并且仅具有睾丸,但是雌性是XX并且具有卵睾,即功能性卵巢和不同数量的非功能性睾丸组织。在他们的第一个模型中,McVean&Hurst(121)考虑了使XX胚胎雄性化Y连锁因子(在我们的术语中,一个父系效应性别决定基因)的进化,这种因子被常染色体或X-染色体上的修饰物抵消。在他们的第二个模型中,他们认为间性体XX个体是雌性驱动X染色体雄性化作用以及恢复功能性雌性的常染色体修饰因子之间平衡的结果。他们的第三种模型是在雄性中驱动Y染色体,其被X连锁抑制因子抵消:当以纯合状态存在时导致部分不育,随后常染色体修饰物入侵在XX间性体中恢复生育能力。我们同意McVean&Hurst(121)的观点,即目前很少有这些模型的经验证据。 结论 遗传冲突在性别决定系统进化中的作用证据不断增长,但仍然是间接的。遗传冲突理论与观察到的多样性一致,包括性染色体驱动系统、动物细胞质性比失调因子和植物中的CMS。 已经为特定系统发展了合理的场景。然而,仅在少数情况下,存在有说服力的证据证明遗传冲突作用,特别是植物中A. vulgare 和CMS的性别决定的遗传多样性。在许多系统中,遗传冲突的调用作用是推测出来的,需要进行进一步实证研究。进一步的理论研究还有很大的空间。 关于遗传冲突和性别决定系统进化的有趣问题包括:( a )X:A平衡系统如何从主要的性别决定基因系统进化以及是否涉及遗传冲突;( b )性染色体驱动和细胞质性别比例失调引起合子性别决定机制的补偿性变化;以及( c )亲本基因/合子基因冲突是否在性别决定进化中发挥作用。我们认为遗传冲突最终将被证明是塑造性别决定机制的重要力量,但这尚待证实。 致谢 特别感谢U Nur多年来关于性别决定和性别比例进化的许多有趣讨论。我们还要感谢D Charlesworth和P Samitou-Laprade对CMS的进化以及D Bopp和ADübendorfer对 Musca 性别决定系统的有益综述和讨论。J Bull关于性别决定进化的开创性工作在我们对这一主题的思考进化中发挥了重要作用。该综述是在JHW在德国Seewiesen的Max-Planck行为生理学研究所的N Michiels研究小组期间开始的,该研究由Alexander von Humboldt-Stiftung的研究奖获得。美国NSF也提供对JHW的支持。荷兰皇家艺术与科学学院的奖学金使得LWB的研究成为可能。 https://blog.sciencenet.cn/blog-536560-1155280.html 上一篇:低覆盖度基因组测序获取系统发育组学数据下一篇:Journal Club:昆虫另类遗传系统进化(Benjamin B. Normark,2003) 收藏 IP: 222.74.29.*| 热度| |
【本文地址】