冠状病毒如何感染我们的细胞|《自然》详解 |
您所在的位置:网站首页 › 你知道新型冠状病毒吗英文怎么说呢 › 冠状病毒如何感染我们的细胞|《自然》详解 |
冠状病毒如何感染我们的细胞|《自然》详解
|
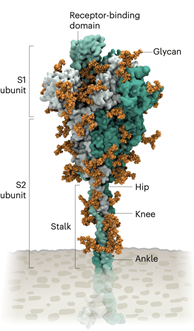
翻译 左文婷 校对 邹晓辉 冠状病毒有一层奢华的糖衣。Rommie Amaro盯着电脑上模拟出来的SARS-CoV-2标志性刺突蛋白——S 蛋白,令人印象特别深刻。这种蛋白从病毒表面伸出,被多聚糖包裹着。加州大学圣地亚哥分校(University of California,San Diego)的计算生物物理化学家阿马罗(Amaro)说:“当它和所有的多糖在一起时,你几乎无法辨认。” 许多病毒的表面蛋白由多糖覆盖,就像披着羊皮的狼一样伪装,逃脱人体免疫系的监视。但去年,阿马罗的团队和合作者基于对蛋白结构和基因数据的了解,用超级计算机在原子水平描绘了这件“糖衣”的结构,进行了迄今为止最精确的可视化。2020年3月22日,她在推特上发布了这一模拟结果。不到一个小时,一名研究人员在评论中问道:从蛋白质顶部伸出的裸露的环是什么? 阿马罗无法回到这个问题。但10分钟后,德克萨斯大学奥斯汀分校(University Of Texas At Austin)的结构生物学家杰森·麦克莱伦(Jason McLellan)解释道:这个未被包被的环是S蛋白的受体结合域 (RBD),是S蛋白三个部位中与人细胞受体结合的区域(图1)。 在Amaro的模拟中,当RBD上移到S蛋白顶端时,两个多糖分子牢牢嵌入到RBD中,就像自行车上的支架一样将其锁定在原地。当Amaro模拟将这两个多糖突变时,RBD结构将崩塌。McLellan的团队用另一种方式在实验室中尝试相同的实验,2020年6月,他们的研究显示将这两个多糖的突变会降低了刺突蛋白与人体细胞受体结合的能力。McLellan说,这一作用在以往对冠状病毒的研究中未被认识到。Amaro认为,去掉这两种糖有可能降低病毒的传染性,尽管研究人员还没有办法证实这一点。 自从新冠肺炎疫情爆发以来,科学家们一直在研究SARS-CoV-2是如何感染细胞的。通过剖析感染过程,他们希望找到更好的方法,通过改进治疗方法和疫苗来阻断感染过程,搞清楚为什么最近的变异株如Delta变异株,更具传染性。 在几十年的冠状病毒研究成果的背景下,经过19个月的研究,我们对SARS-CoV-2是如何入侵人体细胞有了详细的认识(详见图2 “新冠病毒的生命周期”)。科学家们已经发现病毒以惊人的力量入侵人体细胞并迅速隐藏起来的关键机制。并且,当病毒离开细胞时,又执行了一个关键的步骤,使其病毒颗粒感染更多的人体细胞。正是这些方法使病毒得以如此迅速地传播,夺走了数百万人的生命。伦敦帝国理工学院的病毒学家温迪·巴克利说:“这就是为什么它如此难控制的原因。” 图1 SARS-CoV-2刺突蛋白的结构。 刺突蛋白将自己包裹在多糖中,躲过免疫系统的识别。 茎部的三个铰链结构,使它具有灵活性。 从刺突蛋白开始 感染过程从刺突蛋白开始。每个SARS-CoV-2病毒颗粒的外表面都分布着24-40个随机排列的刺突蛋白,这是它与人类细胞融合的关键。对于其他类型的病毒,如流感病毒,表面融合蛋白相对刚性不够灵活。然而,根据德国法兰克福马克斯普朗克生物物理研究所的生物化学家马丁·贝克(Martin Beck)和他的同事在2020年8月的研究成果,SARS-CoV-2病毒的刺突蛋白具有3个像铰链一样的结构,使SARS-CoV-2刺突蛋白具有极大的灵活性(图1)。 这种结构使刺突蛋白可以掉头、摆动和旋转,更容易扫描细胞表面,使多个刺突更容易与人类细胞结合。贝克说,其他冠状病毒没有类似的实验数据,但由于刺突蛋白序列在进化上高度保守,可以合理地假设这一特征是共有的。 在大流行早期,研究人员证实,SARS-CoV-2刺突蛋白的RBD附着在一种名为ACE2受体的蛋白质上,这种蛋白质位于人类咽喉和肺脏细胞的表面。该受体也是导致严重急性呼吸综合征(SARS)的SARS-CoV病毒进入细胞的位置。但与SARS-CoV相比,SARS-CoV-2与ACE2受体的结合强度估计是其2-4倍,其RBD区域的几个变异稳定了病毒与细胞结合的热点区域。危险的SARS-CoV-2变异株往往在刺突蛋白的S1亚基发生突变, RBDS在该亚基上面并负责与宿主ACE2受体结合(刺突蛋白的第二个亚基S2促使病毒与宿主细胞膜融合)。 例如,Alpha变异株刺突蛋白上发生了10个变异,这导致RBD更有可能处于“靠上”的位置。北卡罗来纳州达勒姆杜克人类疫苗研究所(Duke Human Vaccine Institute)的一位研究刺突蛋白突变的结构生物学家普里亚姆瓦达·阿查里亚 (Priyamvada Acharya)认为:“这些变异使病毒更容易进入细胞,从而帮助病毒传播。” 目前正在世界各地传播的Delta变异株在S1亚基中有多个突变,其中包括RBD中的三个突变,这似乎可以提高RBD与ACE2亲和力和逃逸免疫系统的能力。
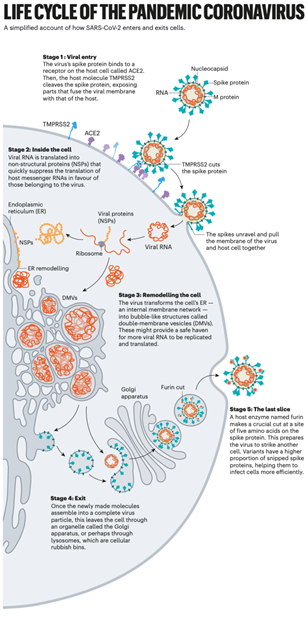
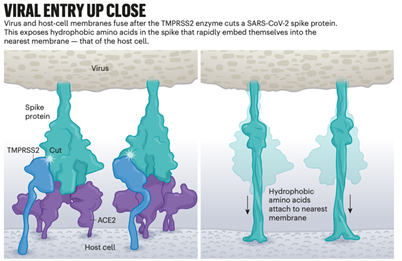
图2 SARS-CoV-2的生命周期 阶段一:病毒进入。病毒的刺突蛋白与宿主细胞上ACE2 受体结合。然后,宿主分子 TMPRSS2 切割刺突蛋白,暴露病毒膜与宿主膜融合的部分。阶段二:胞内。病毒RNA被翻译成非结构蛋白 (NSP),可快速抑制宿主mRNA的翻译,以支持病毒mRNA的翻译。阶段三:重塑细胞。病毒将细胞的内质网(一种内部膜网络)转化为称为双膜囊泡 (DMV) 的气泡状结构。这可能为更多病毒 RNA 的复制和翻译提供了一个安全的避风港。阶段四:出胞。一旦新制造的分子组装成完整的病毒颗粒,就会通过高尔基体或通过溶酶体离开细胞。阶段五:最后一步。一种名为Furin的宿主酶在刺突蛋白上的五个氨基酸的位置上进行了关键的切割。这就为病毒攻击另一个细胞做好了准备。变异体具有更高比例的被切割的刺突蛋白,这有助于它们更有效地感染细胞。 限制性的入口 一旦病毒刺突蛋白与ACE2结合,宿主细胞表面的其他蛋白质就会启动病毒和细胞膜的融合过程。 SARS-Co使用两种宿主蛋白酶中的任何一种来入侵:TMPRSS2 (读作Tempress Two)或组织蛋白酶L。TMPRSS2是较快的入胞途径,但SARS-CoV通常通过内体进入细胞,内体是一种脂质包围的囊泡,这个途径依赖组织蛋白酶L。然而,当病毒颗粒通过这种途径进入细胞时,抗病毒蛋白可以捕获它们。 SARS-CoV-2与SARS-CoV的不同之处在于它有效地利用了TMPRSS2,TMPRSS2是一种大量存在于呼吸道上皮细胞表面的酶。TMPRSS2首先在刺突蛋白的S2亚基上进行切割,这个切割会暴露了一系列疏水氨基酸,这些氨基酸迅速嵌入距离自己最近的膜—宿主细胞膜中。接下来,延长的刺突会像拉链一样折叠起来,迫使病毒和细胞膜融合 (参见图3,“病毒近距离进入”)。 然后,病毒将其基因组注入到细胞内。根据巴克莱和她在伦敦帝国理工学院的同事4月份发表的研究,通过这种弹簧式的入侵方式,SARS-CoV-2比SARS-CoV感染得更快,并避免被困在内体中。 SARS-CoV-2通过TMPRSS2途径快速入侵细胞解释了为什么治疗疟疾的药物氯喹,尽管在实验室的早期研究中很有希望,但在临床试验中表现不好,因为氯喹只作用于依赖组织蛋白酶通过内体方式入胞的病毒。巴克莱说:“当病毒在人的气道中传播和复制时,它不通过核内体,而氯喹是一种干扰核内体的药物,所以在现实中用于治疗新冠是无效的”。 这一发现还指出,蛋白酶抑制剂是一种有希望的治疗选择,可以防止病毒使用TMPRSS2、组织蛋白酶L或其他蛋白酶进入宿主细胞。日本批准用于治疗胰腺炎的TMPRSS2抑制剂-甲磺酸卡莫司他阻止了病毒进入肺细胞,但在最初的临床试验中,该药物并没有改善患者的预后。 德国哥廷根灵长类中心感染生物学部门主任斯特凡·波尔曼(Stefan Pöhlmann)说:“在我看来,我们在一开始就应该用蛋白酶抑制剂作为广谱抗病毒药物来阻止疫情暴发和发展为后来的全球大流行”。斯特凡领导了ACE2结合位点和TMPRSS2途径的研究。
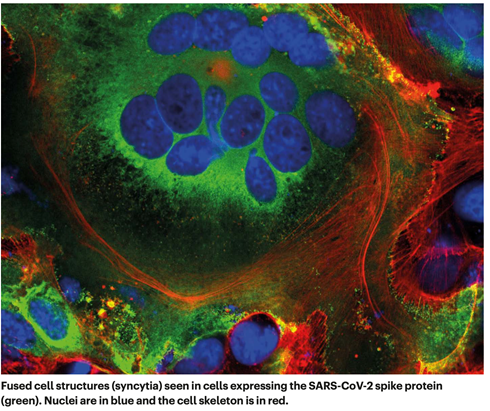
图3 SARS-CoV-2入胞近景 致命的竞争 接下来的感染步骤就比较模糊。盐湖城犹他大学的化学家珍妮特·伊瓦萨正在制作一个带注释的病毒生命周期动画,她说:“一旦病毒进入细胞内就会发现很多黑箱,更多的是不确定性,以及相互竞争的假设。” 在病毒将其RNA基因组射入细胞后,细胞质中的核糖体将病毒RNA的两个部分翻译成长链氨基酸,然后被剪断成16种蛋白质,其中包括许多参与RNA合成的蛋白质。随后,产生了更多的RNA,这些RNA编码了总共26种已知病毒蛋白,包括用于制造新病毒颗粒(如刺突蛋白)的结构蛋白,以及其他辅助蛋白。通过这种方式,病毒开始大量复制自己的mRNA。但它需要细胞翻译机器来将这些mRNA翻译成蛋白质。 冠状病毒在许多方面接管了这一机制。病毒学家诺姆·斯特恩·吉诺萨(Noam Stern Ginossar)和她在以色列雷霍沃特魏茨曼科学研究所的研究小组深入研究了SARS-CoV-2抑制宿主mRNA翻译的三种机制。斯特恩·吉诺萨说,没有一种机制是这种病毒所独有的,但这些作用的组合、速度和程度似乎是独一无二的。 首先,病毒消除了竞争:病毒蛋白Nsp1是病毒进入细胞时最先翻译的蛋白质之一,它招募宿主蛋白系统的切割所有没有病毒标签的细胞mRNA。当Stern-Ginossar的研究小组在宿主mRNA的末端贴上同样的标签时,该mRNA没有被切割。 其次,感染会使细胞内蛋白质的整体翻译能力减少70%。根据两个研究小组的研究,Nsp1再次成为罪魁祸首,这一次是物理上阻断了核糖体的进入通道,使mRNA无法进入。Stern-Ginossar说,剩下的少量翻译能力专门用来翻译病毒的RNA。 最后,病毒关闭了细胞的警报系统。这存在多种机制,但Stern-Ginossar的团队发现了SARS-CoV-2的一种明确机制:该病毒阻止细胞mRNA流出细胞核,包括旨在提醒免疫系统注意感染的蛋白质的指令。第二个研究小组证实了这一发现,并再次指向Nsp1:这种蛋白质似乎堵塞了细胞核中的离核通道,因此没有东西可以离开。 因为基因转录物不能离开细胞核,所以受感染的细胞不会释放太多干扰素——干扰素是提醒免疫系统注意病毒存在的信号蛋白。SARS-CoV-2在关闭这一警报系统方面特别高效:与包括SARS-CoV和呼吸道合胞病毒在内的其他呼吸道病毒相比,SARS-CoV-2感染诱导的干扰素水平明显较低。今年6月,研究人员报告了Alpha变异株的突变似乎使其能够更有效地抑制干扰素的产生。 Stern-Ginossar说:“很明显,SARS-CoV-2有一种独特的能力,可以在第一阶段阻止我们的免疫系统识别和抗击感染”。当免疫系统意识到有病毒存在时,病毒的数量已经太多了,以至于免疫应答蛋白有时会比正常速度更快的速度涌入血液,从而造成机体损伤。医生们在疫情大流行的早期就发现,一些新冠肺炎携带者病情严重,除了病毒本身外,对SARS-CoV-2过度激活的免疫应答也会对他们造成伤害。一些已被证实有效的治疗方法正是通过抑制这种免疫反应起作用。 改造站 一旦病毒接管了宿主的翻译,它就会开始改头换面,根据需要对细胞的内部和外部进行广泛的改造。 首先,一些新制造的病毒刺突蛋白转运到细胞表面并突出宿主细胞膜外。在那里,它们激活宿主钙离子通道,在细胞表面形成一层脂质层——与自然融合的细胞(如肌肉细胞)上的脂质层相同。此时,受感染细胞与表达ACE2受体的相邻细胞融合,发育成充满多达20个细胞核的巨大单个呼吸道上皮细胞(图4)。 伦敦国王学院(King‘s College London)的分子生物学家毛罗·吉亚卡(Mauro Giacca)表示,这些融合结构被称为合胞体,HIV和单纯疱疹病毒等病毒感染可以诱导产生合胞体,但SARS病毒不会。他领导的团队在4月18日发表了这一发现。他推测合胞体的形成可以让受感染的细胞在很长一段时间内茁壮成长,大量生产越来越多的病毒粒子。他说,“这不是一种肇事后会逃跑病毒,他会持续存在下去”。 由北京的中国医学科学院研究员孙强领导的另一个研究小组发现,一些SARS-CoV-2感染细胞甚至与淋巴细胞(人体自身的免疫细胞之一)形成合胞体。这是已知的肿瘤细胞免疫逃避机制,但不是病毒的机制。这表明,被感染的细胞通过简单地抓住并融合附近的免疫侦察细胞来逃避免疫监视。 在细胞内部发生了更多的变化。像其他冠状病毒一样,SARS-CoV-2将细长的内质网(ER),转变为双层膜对囊泡,内质网是参与蛋白质合成和运输的扁平膜网络结构,就像内质网在吹泡泡一样。这些双膜囊泡(DMV)可能会为病毒RNA的复制和翻译提供一个安全的场所,保护它们免受细胞内的天然免疫传感器的影响,但这一假设有待进一步的验证。 参与制造DMV的蛋白质可能是很好的药物靶点,因为它们似乎是病毒复制所必需的。比如,宿主蛋白TMEM41B需要被用来动员胆固醇和其他脂质来扩大ER膜,以便所有病毒部分都能进入。德州大学加尔维斯顿分校的冠状病毒研究员维尼特·梅纳克里(Vineet Menachery)参与了这项研究,他说:“当取走TMEM41B时,会对感染过程产生重大影响。”冠状病毒跨膜蛋白NSP3也可能成为靶点:它可以在DMV的壁上形成一个冠状的孔,以便新产生的病毒RNA穿梭出来。 大多数病毒都有一个外部包装,也就是所谓的包膜,它们直接在细胞边缘组装,在离开细胞的过程中利用细胞自身的质膜来形成这种特征。但新合成的冠状病毒蛋白质走了一条不同的道路。 多年来,一直有证据表明,冠状病毒是通过高尔基复合体(Golgi Complex)运出细胞的,高尔基复合体是一种细胞器,其功能类似于邮局,将分子包装在膜上,然后将它们运送到细胞的其他部分。马里兰州巴尔的摩约翰霍普金斯大学的病毒学家和细胞生物学家卡罗琳·马查默(Carolyn Machamer)研究冠状病毒已有30年之久,她说,病毒从高尔基复合体的膜上形成脂质包膜;然后,新形成的病毒粒子在高尔基囊泡内被带到细胞表面,在那里它们被吐出细胞。 但在12月,位于马里兰州贝塞斯达的美国国家心脏、肺和血液研究所的细胞生物学家Nihal Altan-Bonnet和她的同事报告说,他们检测到冠状病毒通过溶酶体离开细胞——溶酶体是一种充满酶的细胞垃圾桶,可以分解细胞。Altan-Bonnet说,阻断基于高尔基体的分泌途径似乎并没有影响感染性病毒粒子的释放。她的研究小组的证据表明,病毒蛋白质通过内质网出芽形成一层包膜,然后占据溶酶体离开细胞。研究人员目前正在测试阻断溶酶体出胞过程的抑制剂,作为潜在的抗病毒候选药物。 与从质膜中出胞相比,通过高尔基体或溶酶体离开细胞是缓慢和低效的,所以科学家们不知道SARS-CoV-2为什么会这样做。马查默怀疑,高尔基体或溶酶体来源的包膜中的脂质成分比质膜来源的脂质在某种程度上对病毒更有利。她说:“如果我们能更好地理解这一点,就会有很好的机会开发新的抗病毒疗法。”
图4. 表达新冠刺突蛋白的细胞形成的融合细胞结构 最后一步 在离开细胞的过程中,最后一个事件使这种病毒成为一种具有传染巨兽:在含有五种氨基酸的位点快速剪切,使病毒准备攻击下一个目标。 其他冠状病毒在刺突蛋白的S1和S2亚基的交界处只有一个氨基酸(精氨酸),而SARS-CoV-2有五个氨基酸组成的序列:脯氨酸、精氨酸、精氨酸、丙氨酸和精氨酸。波尔曼说:“这个部位不同寻常,所以我们把重点放在了此处,结果证明,这个部位对于肺脏细胞的入侵是必不可少的。”2020年5月,他和他的同事报道说,一种名为Furin的宿主细胞蛋白识别并切割了这一串氨基酸—而这种切割对病毒有效地进入人类肺脏细胞是“必不可少的”。巴克莱说,这不是研究人员第一次在病毒上发现Furin蛋白裂解位点;高致病性禽流感病毒也有这种位点。当一位同事给巴克莱送去一种天然失去Furin蛋白裂解位点的SARS-CoV-2毒株时,她的研究小组发现,感染这种毒株的雪貂比感染大流行毒株的雪貂分泌的病毒颗粒更少,而且不会感染附近的动物。就在巴克莱的研究小组在2020年9月的预印本上报告其结果的同时,荷兰的一项研究还发现,具有完整Furin裂解位点的冠状病毒比没有该位点的冠状病毒进入人类气道细胞的速度更快。 有人怀疑,在病毒粒子组装过程中或者在释放之前的某个时刻,Furin会切割该部位。伊利诺斯芝加哥洛约拉大学病毒学家Tom Gallagher说,这可能解释了为什么病毒会通过高尔基体或溶酶体排出体外。“病毒一旦组装完成,就会进入一个细胞器,在那里有Furin蛋白酶的存在”, Gallagher这样说。Furin通过切断S1和S2亚单位之间的键使病毒刺突蛋白松动,从而在进入细胞时,对TMPRSS2的第二次切割作出反应,TMPRSS2暴露了疏水区域,该区域迅速插入宿主细胞膜。 如果刺突没有被furin预先剪断——而且它们并不总是这样——它们就会绕过TMPRSS2,并通过较慢的内体途径进入,或者根本进不去。 新冠病毒Alpha 和Delta 变异株furin裂解位点都发生了变化。在Alpha变异株中,原来脯氨酸变为组氨酸(P681H);在Delta变体中,它变为精氨酸(P681R)。Barclay说,这两种变化都使氨基酸序列的酸性降低,氨基酸序列碱性越高,furin识别和切割它的效率就越高。”。我们可以假设,这使病毒获得了更好的传播能力。更多的furin切割意味着更多的刺突蛋白进入人类细胞。梅纳赫里说,在SARS病毒中,只有不到10%的刺突蛋白被启动,他的实验室小组一直在量化启动的刺突蛋白,但尚未发表这项工作。在SARS-CoV-2中,这一比例上升到50%。在Alpha变异株中,超过50%。该研究小组发现,在高度可传播的Delta变异株中,超过75%的刺突是预先准备好感染人类细胞的。 更多的未知 科学界对SARS-CoV-2的理解仍然停留在皮毛上。McLellan说,关键的未知领域包括:与每个刺突蛋白结合所需的ACE2受体的数量;TMPRSS2何时切割S2位点;病毒-细胞膜融合所需的刺突蛋白的数量——这只是入胞过程中的未知。2020年4月,加州大学旧金山分校的一个研究小组确定了SARS-CoV-2与人类蛋白质之间至少332个相互作用。 跟上病毒快速变异的步伐并不容易。专家们一致认为,迄今为止,大多数突变都与病毒传播的效率有关,而不是病毒对宿主的破坏程度有关。本月,一项研究报告称,与早期版本的病毒相比,Delta 变体在人体肺部和咽喉的生长速度更快,水平更高。 但尚不确定 Delta 的突变是如何以这种方式增强变异的。Stern-Ginossar 说,“这是许多实验室试图弄清楚的问题”。 |
【本文地址】
今日新闻 |
推荐新闻 |