梨S基因与S基因型鉴定的研究进展 |
您所在的位置:网站首页 › STR基因座命名 › 梨S基因与S基因型鉴定的研究进展 |
梨S基因与S基因型鉴定的研究进展
|
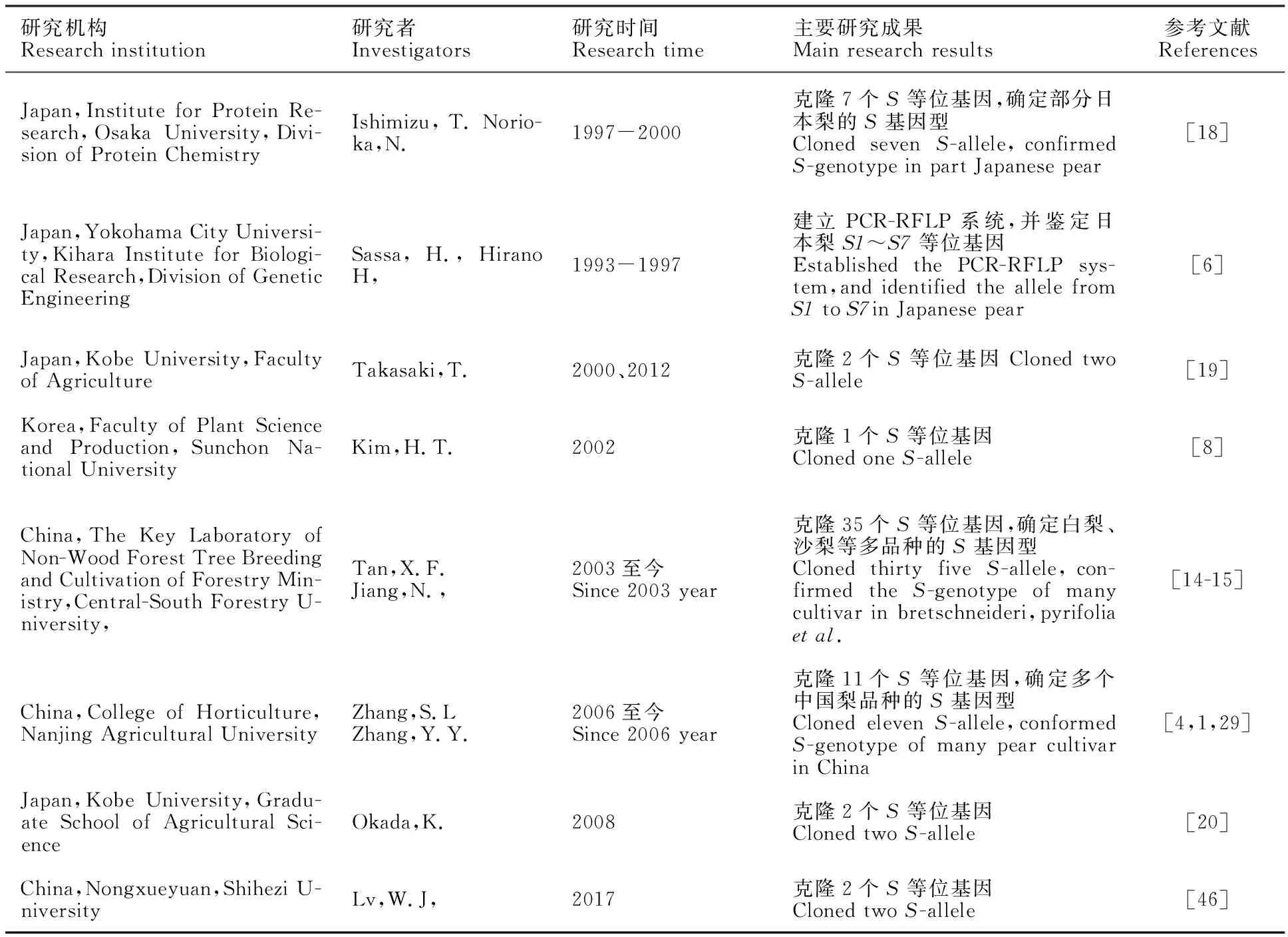
张校立,艾沙江·买买提,徐叶挺,邓 莉,王继勋 (新疆农业科学院 园艺作物研究所/农业部新疆地区果树科学观测试验站,乌鲁木齐 830091) 植物自交不亲和性(Self-incompatibility,SI)是指能产生具有正常功能且同期成熟的雌雄配子的雌雄同体植物,在自花授粉或相同基因型异花授粉时不能完成受精的现象。自交不亲和现象普遍存在于绿色开花植物中。对植物自交不亲和现象的研究始于20世纪30年代,到目前为止己对蔷薇科、茄科、十字花科、菊科等70多个科250个属的绿色开花植物开展了自交不亲和性研究[1]。 梨在植物分类上属于蔷薇科苹果亚科梨属,是由单一位点的S等位基因控制的典型配子体自交不亲和性植物,其雌蕊花柱内的S基因产物为具有核酸酶(RNase)活性的糖蛋白S核酸酶(S-RNase),特异地控制花粉和雌蕊识别过程[2],生产中须合理配置授粉树或采用人工授粉等辅助措施才能保证坐果。中国梨品种资源丰富,有3 000多个品种资源,蕴含有丰富的S基因资源[3];因此,鉴定梨品种S基因型、挖掘梨S基因资源对梨丰产栽培与遗传改良育种具有重要意义。 1 梨S基因型鉴定研究进展早期对梨S等位基因命名方式有两种,即阿拉伯数字与英文字母命名法,其中,亚洲梨种群使用阿拉伯数字命名,西洋梨种群使用英文字母命名。随着对S基因的深入研究,一些S基因被发现同时存在于中国古老的品种或野生类型个体和西洋梨品种中[4]。于是,国外的学者对西洋梨种群的S基因以阿拉伯数字命名的方式重新作了梳理和命名[5]。 梨自交不亲和性基因的研究始于20世纪50年代。国外,最早是Kikuchi(1929年)、Machida等(1982年)和Terami等(1946年)日本学者,采用广泛的田间授粉杂交试验,从日本梨中鉴定7个S等位基因,命名 S1、S2、S3、S4、S5、S6和 S7。Sassa等[6]于1999年首先对 S1~ S7 RNase的cDNA进行克隆,建立日本梨 S1~ S7等位基因PCR-RFLP系统。2000至2001年,Castillo等[7]使用该系统,发现 S8和 S9 2个新的S等位基。2006年,Kim等[8]利用PCR-RFLP又在梨品种‘Chengsilri’中鉴定出 S10-RNase新基因。在西洋梨中,Zisovich 等[9]、Takasaki 等[10]与Mota等[11]利用特异引物和基因克隆的方法鉴定出Sa~St(Sa,Sb,Sc,Sd,Se/Sj,Sh,Si,Sk,Sl,Sm,Sn,So,Sp,Sq,Sr,Ss和St)等17个S-RNaseS基因。2009年,Javier[12]在西洋梨品种‘Abugo’ 和 ‘Ceremeno’中,鉴定出1个新的自交亲和的S-allele: S21。这个基因是 S21的突变体。2014年,Azamnikzad等[13]在57个伊朗梨品种中鉴定出3个新的S-RNase,分别是 S126、S127 和 S128 。国内,最早是乌云塔娜[14]在2003年,从中国白梨中分离鉴定7个新的S基因,分别记为 S17、S19、 S20、S21、S22、S26和 S27等位基因;Tan等[15]分别从白梨、沙梨及秋子梨等亚洲梨种群分离鉴定34个S等位基因;据文献[16]报道,张绍铃等也从亚洲梨种群分离鉴定11个S等位基因,至今为止,东方梨中己获得59个S-RNase等位基因登录GenBank(表1),其中47个等位基因序列完善,并确定部分梨品种基因型,从西洋梨中己鉴定53个等位基因均己登陆GenBank。另外,武军凯[17]研究发现,‘金坠梨’自交亲和性突变不是由花粉S因子突变导致的,而是非S位点的因子突变导致的,这是首次在蔷薇科苹果亚科中发现了由非S因子突变导致的自交亲和性突变体,为蔷薇科自交不亲和机制的研究提供重要依据; 为更好地掌握梨S等位基因和S基因型鉴定方法及研究过程,将已确定的500多个梨品种S基因型进行整理和汇总(表2),并对各科研单位的研究成果列表(表3)展示,以期为梨S等位基因深入研究和梨树丰产栽培提供指导。
表1 梨栽培品种 S 等位基因命名Table 1 Numbering of S-alleles in pear cultivars
表2 梨S等位基因及 S基因型鉴定研究现状Table 2 Summary of the research of S-allele and S-genotyping in pear 2 梨自交不亲和S基因型的鉴定方法早期日本学者采用田间授粉试验等传统方法对梨自交不亲和S基因型进行鉴定,利用梨相同S基因型品种间杂交不亲和性的原理,进行大量杂交试验,鉴定7个S基因,确定40多种日本梨品种的S基因型[21],此方法的优点是简单易操作,缺点是工作量大、鉴定周期长、效率低,受生理和环境条件的影响大,结果可靠性差,当双亲或一个亲本的S基因型未知或没有适当的授粉测试品种时,难以用该方法确定品种的S基因型。授粉花柱离体培养法的优点是省工、高效,缺点是试验操作技能要求高。随着生物技术的不断发展,S基因型的鉴定方法也有了很大发展,PCR-RFLP技术[6]、蛋白质电泳分析[22]、DNA序列分析方法[23]和基因芯片杂交[24]等技术均被用于梨品种自交不亲和S基因的研究中,蛋白质电泳分析法具有快捷 、可靠等优点,缺点是操作过程较复杂,操作技术要求较高;DNA序列分析方法具有简单、准确、效率高等优点,缺点是成本高,不能独自鉴定S基因,需要与PCR扩增技术等方法联合使用。PCR-RFLP技术具有快捷、准确、效率高,操作简便等特点,并且试验材料取样简便,缺点是难以对含有新S基因的品种的基因型确定,需结合S基因特异序列DNA测序方法等其他鉴定方法;基因芯片杂交技术具有简单,快速,准确,高效,灵敏度高,可实现对多个梨品种的S基因型同时检测,并具有大量样品平行检测的优势。缺点是具有一定的局限性,只能检测出已发现的梨S基因,不能鉴定出新的S基因,需结合S基因特异序列DNA测序方法等其他鉴定方法。目前,这些S基因型的鉴定方法以PCR-RFLP技术和S基因DNA序列分析法应用较为广泛,梨S基因芯片杂交技术是近年来发展起来的新的鉴定方法,拥有广阔的应用前景。 2.1 S基因特异PCR技术在研究早期,日本学者通过对已知的7个S基因的DNA及cDNA序列比较发现,梨的S基因具有5个保守区、1个高变区(hypervariable region,HV)及1个内含子区,其HV区是在自交不亲和反应中特异识别花粉的专一功能区,且不同S基因间HV区的多态性很高[25-26]。在此基础上1999年Sassa等[6]建立梨 S1~S7等位基因PCR-RFLP系统,即根据S基因中保守序列设计引物,对基因组DNA进行PCR扩增,用S基因特异的限制性内切酶消化扩增片段,由于高变区和内含子序列具有S特异性,故酶切片段具有长度多态性,通过电泳与己知S基因酶切片段大小比较,即可确定梨品种的S基因型[27]。2000-2001年,Castillo[28]使用 S1~S7等位基因PCR-RFLP系统时,发现 S8-和 S9-两个新的S等位基因,并对其氨基酸序列进行研究,建立 S1-9等位基因PCR-RFLP系统。
表3 梨品种S基因型总结Table 3 Summary of S-genotypes of pear cultivars
(续表3 Continued table 3)
(续表3 Continued table 3)
(续表3 Continued table 3) PCR-RFLP技术相较于田间授粉试验、蛋白质电泳分析等方法,具有速度快、准确高、取样简便等优点,但其是在已获得S基因且基因序列已被分析的基础上进行鉴定,难以确定含有新S基因品种的基因型,需结合S基因特异序列DNA测序等其他鉴定方法。 DNA测序方法是对基因组DNA的PCR扩增产物进行回收测序,然后在GenBank进行BLAST以确定梨品种的S基因型[29],分析新的自交不亲和S基因。此方法不仅快捷、准确、操作简便,且试验材料不受季节限制,大大加快试验进程。谭晓风等(2005年)对中国沙梨S基因进行PCR-RFLP分析,对采用PCR-RFLP方法无法确定的S等位基因和S基因型进行目的片段的DNA测序,首次从中国沙梨中分离鉴定了7个新的S基因,并确定近20个中国沙梨的S基因型。目前为止,运用此方法Tan等[15]与江南等[24]在中国梨中发现35个新自交不亲和S基因,并且鉴定90多个沙梨品种、15个西洋梨品种和近30个白梨品种的S基因型。 2.2 基因芯片杂交基因芯片(Genechips)又称DNA芯片(DNAchips)或DNA微阵列(DNA microarray)[30],是传统的核酸杂交技术与微加工技术以及化学合成技术相结合而产生的一个复合体,基因芯片杂交技术具有检测方法简单、快速、准确、高效、灵敏度高、可实现对多个梨品种S基因型同时检测及具有大量样品平行检测的优点[31]。江南[32]于2006年首次使用梨S基因寡核苷酸芯片对梨品种S基因型进行鉴定,其检测结果获得成功,证明基因芯片杂交技术应用于梨品种自交不亲和性S-RNase基因和S基因型的检测是一种切实可行的方法。2008年江南等[33]又指出,基因芯片技术只能检测出已发现的梨S基因,不能鉴定出新的S基因,具有一定局限性。对于梨品种中存在的新S基因,还需结合PCR-RFLP、DNA测序及序列分析等技术进行鉴定。并且S基因寡核苷酸芯片技术是采用梨DNA序列进行杂交的,存在内含子序列不同其杂交信号也会有差异的问题,针对此问题,江南等[34]以cDNA序列制作梨S基因芯片进行进一步研究,结果发现两种基因芯片各有利弊,利用两种芯片并行检测梨品种S基因,能保证鉴定结果的准确性和可靠性。近两年,江南等[35-36]及谭慧等[21,37]利用梨S基因芯片技术确定86个梨品种的未知S基因型,结合其他S基因型鉴定技术发现7个新的S-RNase基因,命名为 Pp S53(PyruspyrifoliaS53)、 Pp S54、Pp S55、Pp S56、Pp S57、Pp S58和 Pp S59。 3 展 望随着越来越多的研究者开展梨自交不亲和性的研究,在更多的梨品种S基因型被确定的同时,也出现以下问题: 已鉴定的梨品种的S基因型的准确性有待验证。由于研究单位及研究方法的不同或者是由于同名异物,导致梨品种自交不亲和S基因型存在鉴定结果不同的现象,这一现象极易引起授粉树搭配不合理,从而引起生产上不必要的损失。如:对于‘苹果梨’的S基因型,巩艳明[38]鉴定为S1S17,张妤艳等[16]鉴定为S17S19,许高歌[39]鉴定为S1Sdm;对于‘早酥梨’的S基因型,许高歌[39]鉴定为S1Sd,与巩艳明[38]的研究结果一致,而杨谷良[40]则鉴定为S22S35;特别是对‘库尔勒香梨’研究,目前已有多位研究者对其S基因型进行鉴定,但结果均不完全相同,张妤艳等[41]鉴定为S21S28,谭晓风等[42]认为是S8S28;杨谷良[40]和唐忠建[43]鉴定结果一致都认为是S22S34,Qi等[44]、Heng[45]等和吕文娟等[46]鉴定的‘库尔勒香梨’的S基因型结果一致,均认为是S22S28,并通过RT-PCR鉴定得到证实。 梨S等位基因信息不断丰富,其命名存在相同基因不同编码序列或相同编码序列但命名不同等问题。出现此问题的原因可能是:①早期对S等位基因的鉴定主要针对 HV 区及周边序列部分片段的测序鉴定,这些片段只占S基因全长序列的一小部分,因此需要得到完整的全长序列来进一步区分或重新命名更正这些相似基因。②基因登录过程中忽略内含子而造成同物异名的现象。如:2007年衡伟等[47]对S基因的DNA序列分析确认 S16(AY249431.2)和 S31(DQ072113.1)为相同S基因;2013年徐非凡[29]认为 S12(EU117115)与 S36(DQ417607)为同一基因、 S17(EU249432)与 S34(DQ269500)为同一基因,与吕文娟的研究结果相同;吕文娟等[46]对 S7(AB002143)与 S27(EF643640.1)的编码序列进行比对,发现其完全相同应为同一基因,江南等[34]也发现砂梨 S15和 S38基因编码序列完全相同;杨谷良等[42]推导氨基酸,从而推测 S8-RNase、 S28-RNase和 S34-RNase很可能是同一基因。 尚有大量梨品种的S基因型等待鉴定。中国拥有3 000多个梨品种,目前只有500多个品种的S基因型被鉴定,还有大量的S基因型未被鉴定。这给应用梨S-基因型合理选配授粉树和指导梨园生产带来很大的局限性。 新的鉴定技术急待开发。梨S基因的鉴定技术虽然有了巨大的突破,鉴定方法也更加方便、简单、准确;但是都还具有一定的局限性,有时需要多种方法联合使用才能准确鉴定,多种方法联合使用极大降低鉴定速度,增加鉴定成本。因此,开发新的S基因型鉴定技术,以较低的成本,简单、快速、准确地确定更多梨品种的S基因型,进一步完善梨品种S基因检测系统,仍然是梨自交不亲和性研究及梨生产实践急待解决的重要问题。 猜你喜欢 亲和性基因芯片等位基因 亲子鉴定中Penta E稀有等位基因28的确认1例川北医学院学报(2022年6期)2022-06-24亲子鉴定中男性个体Amelogenin基因座异常1例智慧健康(2021年17期)2021-07-30出生时即可预判发育潜力 基因芯片精准筛选肉牛良种今日农业(2021年4期)2021-06-09广东汉族人群Penta D基因座off-ladder稀有等位基因分析中国产前诊断杂志(电子版)(2020年1期)2020-05-21基因芯片技术在生物研究中的应用进展知识文库(2020年5期)2020-05-11贵州汉族人群23个STR基因座的OL等位基因研究遵义医科大学学报(2020年6期)2020-02-05荔枝高接品种的选择世界热带农业信息(2019年9期)2019-01-054个苹果观赏品系开花、授粉习性及花粉管萌发的荧光显微观察山东农业科学(2017年5期)2017-06-05基于MECA算法的BP网络研究数字技术与应用(2016年10期)2017-04-01德国“儿童大学”理念的发展东方教育(2016年14期)2017-01-16
|





 西北农业学报2018年8期
西北农业学报2018年8期【本文地址】
今日新闻 |
推荐新闻 |