Nature |
您所在的位置:网站首页 › 浮游植物的作用和意义 › Nature |
Nature
|
[快速阅读] 全球光合作用消耗的二氧化碳是人为净排放量的10倍,其中微藻占了将近一半。藻类光合作用的高效性依赖于羧化酶 RuBisCO 催化位点的 CO2聚集机制(CCM),这种机制有助于 CO2的固定。虽然已经确定许多细胞组分参与无机碳的运输和固存,但是微藻如何提供能量使CO2在一个热力学梯度下聚集仍然是未知的。2022年4月27日,法国马赛大学Gilles Peltier等人在Nature(自然)期刊发表了题为“Alternative photosynthesis pathways drive the algal CO2-concentrating mechanism”的文章,这篇文章证明,在绿藻类的莱茵衣藻中,环状电子流和 O2光还原分别依赖于 pgrl1和 flavodiiron 蛋白的联合作用产生了一个对 CCM 功能至关重要的低腔 pH 值。本文报道了发光质子被用于类囊体bestrophin转运体的下游,可能用于碳酸氢盐转化为二氧化碳。本文进一步确定,电子流从叶绿体到线粒体有助于激活非类囊体无机碳转运体,可能通过提供 ATP的途径。本文提出了一个为CCM提供能量的网络的综合观点,并描述了藻类细胞如何从光合作用分配能量来为不同的CCM过程提供能量。这些结果为将功能性藻类CCM转移到植物中以提高作物产量提供了一条途径。  [研究背景] 水生生态系统中的微藻光合作用必须克服CO2在水中缓慢扩散导致的低CO2可用性。固定二氧化碳的酶RuBisCO对二氧化碳的亲和力较低,因此藻类光合作用的效率高度依赖于CCM。藻类CCM涉及位于不同细胞区域的无机碳(Ci)转运蛋白和碳酸酐酶的依次作用,并导致CO2在RuBisCO层面的活跃积累。在绿藻类C.reinhardtii(以下简称衣藻)中已鉴定出几种CCM成分,特别是推测的跨质膜(高光激活3型(HLA3))、叶绿体包膜(低碳诱导A型(LCIA))和类囊体膜(类bestrophin蛋白转运蛋白14型(BSTs))的Ci转运蛋白。Ci最终通过碳酸酐酶15(CAH3)转化为CO2,从而使其能够被RuBisCO固定。Ci在浓度梯度下穿过双层膜的运输及其转化为CO2是能量依赖过程,光合作用在提供CCM功能所需的化学能方面发挥作用。 在光合作用过程中,两个光系统(PSII和PSI)通过线性电子流(LEF)将太阳光转化为化学能,将NADP+还原为NADPH,并在类囊体膜上产生pH梯度。然后利用pH梯度进行ATP合成,NADPH和ATP都为CO2固定提供能量。然而,LEF产生的ATP少于固定CO2所需的ATP,光合作用依赖于其他机制来重新平衡这一比率。这包括(1)围绕PSI的循环电子流(CEF),涉及植物和藻类中的质子梯度调节子5(PGR5)和质子梯度调节类1型(PGRL1)蛋白质;(2)伪循环电子流(PCEF),将电子转移到PSI受体侧的O2,由蓝藻,苔藓植物和绿色微藻中的黄二铁蛋白(FLV)催化。CEF和PCEF在不产生NADPH的情况下产生pH梯度,从而重新平衡LEF的高NADPH/ATP比率。另一种途径涉及叶绿体和线粒体之间的几种代谢穿梭,我们称之为叶绿体到线粒体电子流(CMEF),当CEF缺乏时,也可以为CO2固定提供额外的ATP。在这种情况下,光合作用能量如何传递给不同的Ci转运蛋白,以及如何在不损害CO2固定能力的情况下实现这一点成为了关键问题。 这篇文章通过研究依赖PGRL1的CEF(pgrl1)、PCEF(flvB)和BSTs的衣藻突变体来解决这些问题。可以表明,CCM活性在单个CEF或PCEF突变体中不受影响,但在双突变体中受到严重损害,并提出这些机制协同作用产生的管腔质子浓度增加被BST类囊体Ci转运蛋白下游的CCM机制所利用。我们进一步证实,CMEF参与CCM的功能,很可能是通过向质膜和/或叶绿体膜Ci转运体提供ATP,从而揭示了远离类囊体的运输步骤是如何被驱动的。 FLVs和PGRL1的缺失会削弱Ci的亲和力 为了研究依赖FLV性PCEF和依赖PGRL1性CEF在CCM能量供应中的作用,我们首先测量了衣藻flvB或pgrl1单突变体在不同Ci浓度下的净光合O2产生量。当细胞在空气(400 ppm CO2(低CO2))下生长时,对照野生型(WT)菌株和单个突变体(半饱和常数(K1/2)≈ 10 µM)的Ci亲和力相似,表示功能齐全的CCM(图1a、b、h)。在高CO2(空气中添加3%CO2)条件下,flvB和pgrl1突变体及其各自的亲本菌株对Ci表现出相似的亲和力,K1/2约为100 µM(图1d、e、h)。为了评估FLV和PGRL1之间可能存在的功能冗余,我们通过单个PGRL1和flvB突变体的遗传杂交获得了双突变体(图S1a)。在子代中,我们分离出五个独立的双突变体(pgrl1 flvB-1、2、3、4和5)以及四个独立的对照菌株,显示FLV和pgrl1(WT1–4)的正常积累(图1g,图S1)。我们观察到,在高CO2条件下生长时,这些菌株之间的Ci亲和力没有差异,但在低CO2条件下生长时,双突变体对Ci的亲和力比对照菌株低七倍(图1c,f,h,图S2b,d)。  图1:PGRL1和FLVB的缺失降低了Ci的光合亲和性  图S1:pgrl1+flvB双变异体的获取和表征 CCM中缺陷的突变体通常不能在低CO2条件下正常生长。我们比较了不同CO2浓度、pH值和光照强度下的生长情况。尽管所有菌株在高CO2条件下都表现出相似的生长,但pgrl1+flvB双突变体在低CO2和极低CO2(空气中100 ppm CO2)条件下的生长受到损害(图2a),类似于CCM缺陷突变体cia5中观察到的生长缺陷(图2a,图S3)。在双突变体中观察到的生长缺陷随着光照强度的增加而恶化,但几乎不受pH值的影响(图S3)。通过免疫检测评估,除LCI1外,所有菌株中主要CCM成分的积累相似,LCI1在双突变体中的含量较低(图2b),在单突变体中的含量较小(图S4c)。然而,LCI1敲除突变体的生长在低CO2下不受影响这一事实表明,在pgrl1+flvB双突变体中观察到的较低LCI1水平不是CCM功能失调的原因,而是后果。体内测得的碳酸酐酶活性在低CO2下诱导,在所有菌株中达到相似水平(图S4f)。双突变体和对照菌株表现出相似的最大O2光合产量(图S2a,c)和PSII量子产量(图S4a),以及相似的主要光合复合物水平(图S4b)。因此,依赖PGRL1性CEF和依赖FLV性PCEF有助于CCM的运行,并可以相互弥补,例如单个突变体中缺乏CCM表型。在强光下观察到的pgrl1突变体的生长损伤表明,PCEF的代偿导致代谢失衡,可能会影响长期生长。  图2: 在低CO2环境中,pgrl1+flvB双突变体的生长因为CCM组分的存在受到了损害  图S2:在pgrl1、flvB、pgrl1+flvB 及其各自的对照中,低 CO2或高 CO2下生长所测量的最大净 O2 释放量和 K1/2 值  图S3:pgrl1、flvB、pgrl1+flvB 突变体的生长及其对照菌株  图S4:光合作用和 CCM 成分评估 CCM使用CEF和PCEF产生的腔内H+ 我们假设PGRL1和FLV可能产生额外的反式类囊体pH梯度,为CCM提供能量。对照菌株和单个pgrl1或flvB突变体在光照条件下产生的pH梯度和质子(H+)动力(PMF)与对照菌株相似(图2c,图S5),而pgrl1+flvB双突变体产生pH梯度和PMF的能力受到严重损害(图2c,图S5)。有人提出,在CCM的最后一步中,可以使用鲁米诺质子转化HCO3− 减少二氧化碳排放。因此,为了进一步了解CEF、PCEF和CCM通电之间的联系,我们评估了不同突变株CCM功能期间管腔质子浓度的变化。我们监测了叶绿素荧光非光化学猝灭(NPQ)的快速可逆成分(qE)的水平,该成分最近被确定为管腔pH的敏感可靠探针(图3a),并取决于在低CO2条件下生长的细胞中捕光复合物应激相关3型蛋白(LHCSR3)的积累(图2b,图S4c)。在对照品系中,当Ci水平较低时,NPQ水平最高,并且在Ci注射时在光线下或在光线关闭时在低Ci下迅速可逆地降低(图3b;图S6a、c、e)。我们确认,状态转变(另一个NPQ成分)不会导致CO2依赖性NPQ变化(图S7a-c),并且在低Ci下测得的NPQ被抑制跨类囊体pH梯度的nigericin离子载体完全放松,因此可归因于qE(图S7d-f,h,i)。qE的CO2依赖性在单个突变体中不受影响。然而,在低Ci(图3c,图S6f,h)下,pgrl1+flvB双突变体中没有诱导qE,尽管这些菌株积累了正常数量的LHCSR3(图2b),并在介质酸化时保持了诱导大量qE的能力(图S6i,j),这与pgrl1+flvB双突变体无法产生pH梯度(图2c)相一致。我们从该实验得出结论,CEF和PCEF通过产生低pH管腔来促进CCM,这两种机制在单个突变体中能够相互替代。  图3: CEF、PCEF和CMEF对CCM贡献能量  图S5:电致变色类胡萝卜素吸光度改变测量值和 pmf 大小以及 ΔpH 和 ΔΨ 之间的分配  图S6:叶绿素荧光、NPQ 和 Ci 在 pgrl1、flvB、pgrl1+flvB 和它们各自的对照组中在暗-亮-暗瞬变期间测量数值  图S7:Ci 依赖的 NPQ 变化与状态转换无关,并与反式类囊体 ΔpH 相关 已经提出三种类BST蛋白在类囊体水平上运输Ci。为了深入了解管腔质子浓度在CCM功能中的作用,我们评估了BST敲除菌株14(bsti-1)中NPQ对重复Ci供应的反应动力学。发现bsti-1可根据生长条件积累不同水平的LHCSR3(图S4e)——因此,我们选择了LHCSR3积累接近对照菌株的条件。而对照菌株在每次Ci注射后显示NPQ降低,然后在Ci耗尽后增加(图3d),bsti-1的NPQ几乎没有受到影响(图3e)。我们验证了bsti-1显示可归因于qE的NPQ成分,并且能够产生与对照菌株类似的pH梯度(图S5、7g)。总之,bsti-1能够产生pH梯度和较低pH的管腔,但不能用于累积Ci。 CMEF为远程Ci转运器供电 为了定量了解pgrl1中补偿机制的本质,我们研究了使用18O标记O2测量的光依赖性O2消耗的Ci依赖性。如前所述,对照菌株在低Ci下的O2消耗增加(图S8a,e)。在pgrl1中,O2摄取率高于对照菌株(图S8a-d),这与PCEF对CEF缺陷的补偿一致。摄氧量显著降低,但摄氧量显著降低(图8F,图28)。为了确定线粒体呼吸是否与剩余的光依赖性O2摄取有关,我们使用了两种线粒体呼吸抑制剂,即粘噻唑和水杨酸异羟肟酸(SHAM),它们分别抑制细胞色素bc1复合物和线粒体替代氧化酶。我们表明,flvB在低Ci下测得的剩余光依赖性O2摄取量确实是由于光合作用驱动的线粒体呼吸活动,因此归因于CMEF(图S8k)。  图S8:在 pgrl1、flvB 及其各自对照组中,存在富含 18O 的 O2 的情况下,使用 MIMS 测量的 O2交换率的 Ci 依赖性 然后,我们研究了线粒体在WT菌株中对CCM激活的贡献。而呼吸抑制剂的添加对高CO2生长菌株(K1/2)的最大净O2产生率(Vmax≈ 100 µM)和Ci亲和力没有影响)(图S9d–f,i, m),它将低二氧化碳生长的对照菌株的Ci亲和力降低了一半(K1/2>20 µM)与未经处理的细胞(K1/2 ≈ 10 µM)相比(图3f)。只有当粘噻唑和SHAM联合使用时,才观察到对Ci亲和力的影响(图S10),这表明交替氧化酶和细胞色素bc1电子途径都有助于CCM的激活。值得注意的是,呼吸抑制剂还增加了空气培养的bsti-1突变体的K1/2(图3f,图S9h),从而表明线粒体对CCM通电的贡献在远离类囊体的转运体水平上起作用。不同途径(CEF、PCEF和CMEF)的贡献可从Ci耗尽期间在不同突变体中测得的O2消耗率得出(图S8)。虽然PGRL1依赖性CEF的贡献保持相对恒定,但FLV依赖性PCEF和CMEF的贡献在低Ci时显著增加(图4a)。  图4:藻类细胞中CCM能量网络的拟议机制  图S9:线粒体呼吸抑制剂对净O2光合作用的Ci亲和力的作用 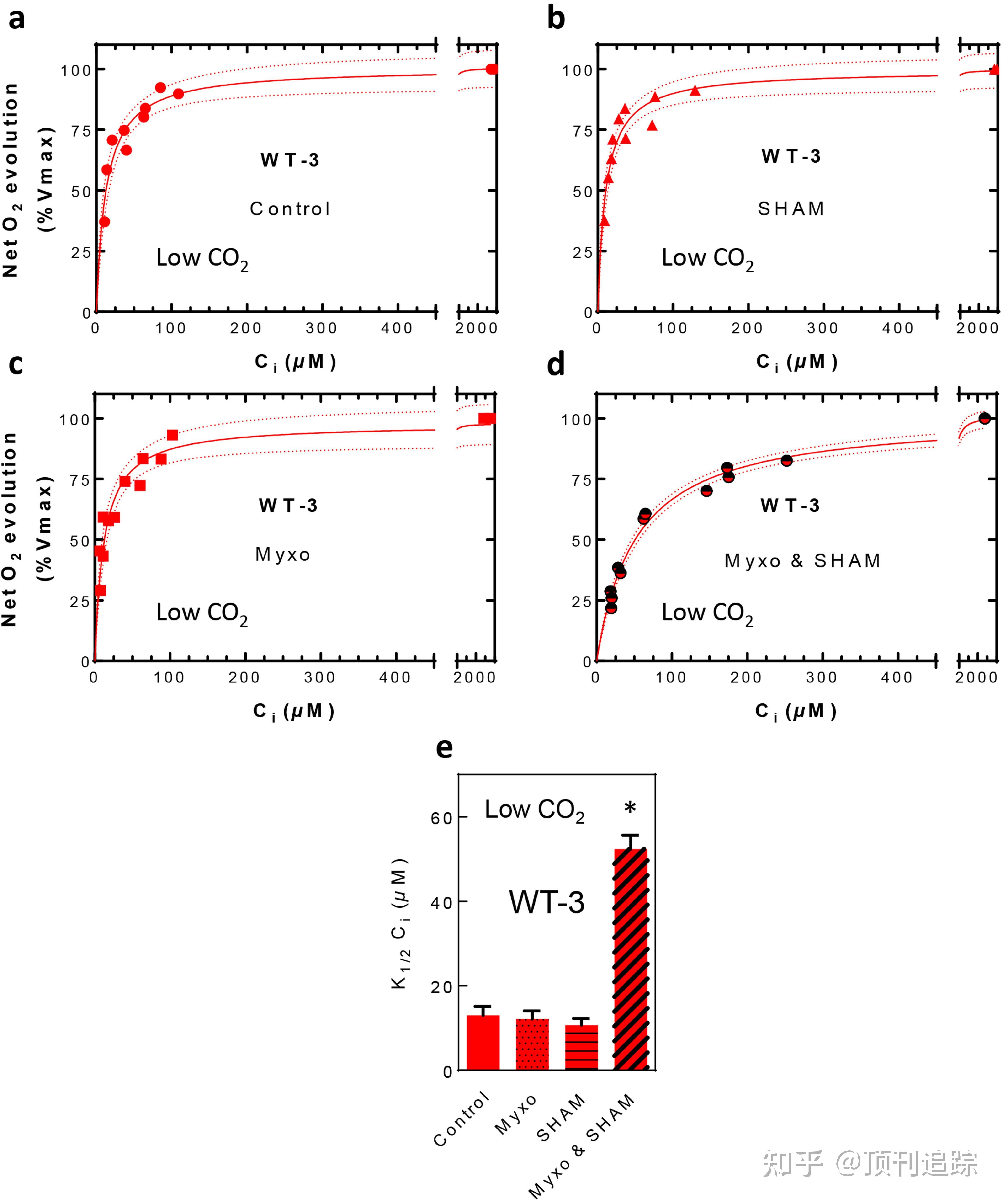 图S10:分别或同时添加呼吸抑制剂myxothiazol和SHAM对净O2光合作用Ci亲和力的影响 尽管CCM对光合作用能量的需求早已被认识到,但相关的分子机制仍然知之甚少。PCEF或CEF的参与已被提出,但其实际和各自的贡献尚未确定。在这里,我们证明了依赖FLV性PCEF和依赖PGRL1性CEF通过产生pH梯度来共同向CCM提供能量。此外,我们提出,这两种机制产生的低管腔pH值被与BST功能相关的类囊体定位CCM过程所利用。因为哺乳动物BSTs38是Cl-对HCO3-具有高度渗透性的通道, 由于CEF和PCEF的联合作用而积聚在管腔中的质子可能用于HCO3−的转化对于由CAH3介导的CO2,这两种机制产生的PMF(图S5)可能有利于HCO3- 通过BST易位(图4b)。 我们进一步证实,线粒体呼吸在CCM活化中的作用在很大程度上被忽视,它通过有效的细胞间氧化还原运输为远离类囊体的CCM转运体提供能量。通过分析每种机制作为Ci浓度函数的各自贡献,我们得出结论,尽管CEF的贡献相对恒定,但PCEF的贡献在低Ci浓度时增加,CMEF的贡献在最低Ci浓度时也变得重要。由于这些是假定的ATP依赖性周质和叶绿体包膜转运体(分别为HLA3和LCIA)高度活跃的典型条件,我们建议CMEF在低Ci下产生的ATP向一个或两个转运体提供能量(图4)。在极低CO2条件下观察到的pgrl1+flvB突变体生长下降较低(图S3d-f)表明,依赖CMEF能量供应的外部转运体(HLA3和/或LCIA)可能是这些条件下CCM的主要驱动因素。值得注意的是,在pgrl1、flvB、pgrl1+flvB和bsti-1中,与HLA紧密相关的LCI1积累减少,表明类囊体水平的CCM损伤可能调节LCI1介导的周质转运过程。这可能是由于缺乏功能性类囊体Ci转运导致细胞溶质Ci浓度增加,进而触发LCI1表达下调,以避免细胞溶质Ci过度积累。 活跃的CCM的存在是影响海洋中浮游植物生物量产生的关键因素,尤其是对产生大量海洋水华的浮游植物物种。然而,由于缺乏对situ9中CCM活动的了解,因此很难预测全球变化将如何影响浮游植物群落。我们在这里证明,功能性CCM的存在可以通过测量依赖Ci的NPQ来探测,NPQ可以用作确定水生生态系统中CCM活性的简单参数。 类囊体CCM过程消耗管腔质子的潜在限制是与二氧化碳固定所需ATP合成的竞争。这一点尤其重要,因为已知LEF提供的ATP少于CO2固定所需的ATP。我们认为,CEF、PCEF和CMEF这三种替代机制的联合作用可以在不影响CO2固定的情况下满足CCM的能量需求,所有这些机制都会导致ATP/NADPH比率的增加。CCM研究中的一个主要生物技术挑战是通过将微藻成分转移到高等植物来提高作物生产力。在植物中建立一个功能齐全的CCM是一个巨大的科学挑战,最近又重新获得了相当大的兴趣。我们的研究表明,对细胞能量学的综合理解是在不影响光合CO2固定效率的情况下满足合成CCM的能量需求的关键。例如,FLVs在高等植物中的表达,已被证明在光合作用期间提供额外的PMF,似乎是提供类囊体Ci运输所需额外能量的一个有希望的起点。我们预计,未来的研究将能源和CCM表达耦合起来,将有助于提高植物生产力。 本公众号承接论文中英互译、论文润色等服务。详情请私信本公众号或发邮件至:[email protected] 欢迎关注公众号:顶刊追踪! 欢迎发私信参与讨论! |
【本文地址】