值得收藏!大肠杆菌基因型及遗传符号说明汇总 |
您所在的位置:网站首页 › 基因型和表现型的意思 › 值得收藏!大肠杆菌基因型及遗传符号说明汇总 |
值得收藏!大肠杆菌基因型及遗传符号说明汇总
|
01 一般规则 ① 根据基因产物或其作用产物的英文名称的第一个字母缩写成 3 个小写斜体字母来表示。 例如:DNA Adenine Methylase→ dam 。 ② 不同的基因座,其中任何一个突变所产生的表型变化可能相同,其表示方法是在 3 个小写斜体字母后加上 一个斜体大写字母来表示区别。 例如:Recombination→ recA、recB、recC。 ③ 突变位点应通过在突变基因符号后加不同数字表示。如 supE44( sup基因座 E 的 44 位突变)。如果不知道几个等位基因中哪一/几个发生了功能性突变,则用连字符“-”代替大写字母,如 trp-31 。 ④ 细菌的基因型中应该包含关于其携带的质粒或附加体的的信息。这些符号包括菌株携带的质粒或附加体、质粒或附加体上的突变基因座和突变位点。 其基因符号应与基因座的表示符号明显区别,符号的第一个字母大写、不斜体并位于括号内;质粒或附加体上的突变基因座和突变位点的基因符号的表示方法与染色体上突变基因座、突变位点的符号相同。 例如: IF' [ traD36、proAB、lac I q、lacZ M15] ⑥ 某个基因或某个领域缺失时,在其基因型前面加上“ ”表示。 例如:lac-proAB 基因缺失时它的基因型表示为 ( lac-proAB)。 ⑦ 由于某种基因的变异导致大肠杆菌可以明显观察到特征变化,有时也用其表现型代替基因型进行表示。 例如:某些抗药性的获得或丧失,用如下方式表示:Streptomycin 抗性→ Str+或 Str r ,Ampicillin 敏感性→ Amp - 。(第一个字母要大写,“+”或“r”表示有抗性,“-”表示无抗性或敏感)。 ⑧ 根据某些特异性蛋白的变异及其导致的结果变化进行表示。 蛋白发生变异,中的 rB-、mB-表示由于 S20 的变异而导致 B 株来源的 hsdR 和 hsdM 的功 能缺失。 ⑨ 蛋白质的名称与对应的基因或等位基因相同,但不用斜体,且首字母大写,如,UvrA、UvrB。 3 基因符号和意义
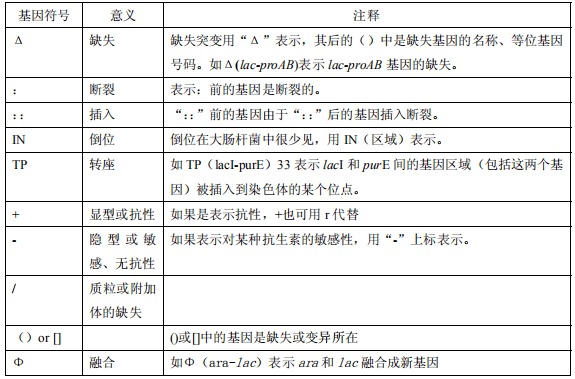
表1 部分基因符号和意义
4 主要的基因型说明 01 基因重组相关的基因型 1 recA (Recombination) 功能:recA 基因表达 ATP 依赖型 DNA 重组酶,它在λ-噬菌体与基因组 DNA 的溶原重组时起作用,同时具有对 DNA 放射性损伤的修复功能。由 recA 基因的变异所产生的基因型使同源或异源 DNA 的重组不能进行,保持插入 DNA 的稳定性,对 DNA 的转化有利。一个菌株的基因型如果是 recA ,则说明此菌株的表现型是重组缺陷的。 2 recB (Recombination) 功能:recB 基因表达 ATP 依赖型 DNase 和核酸外切酶 V 的一个亚基,对 recA 的 DNA 重组酶起辅助和促进作用。DNase 催化双链 DNA 的解旋和解链,核酸外切酶 V 催化单链 DNA的裂解,在 DNA 的重组和损伤修复中发挥重要作用。recB 基因的变异导致其 DNA 重组和修复功能丧失,保证了外源 DNA 的稳定,有利于 DNA 转化。 3 recC (Recombination) 功能:recC 基因表达四种酶,即核酸外切酶 V,ATP 依赖型的核酸内切酶,解旋酶及 ATP酶,它们和 recA, recB 所表达的酶相互协调作用,在 DNA 的重组及放射性损伤的修复中发挥作用。recC 基因的变异导致 DNA 重组功能缺失,保证外源 DNA 的稳定性。 02 甲基化相关的基因型 1 dam (DNA adenine methylase) 功能:dam 基因表达 DNA 腺嘌呤甲基化酶,它能催化特异序列 GATC 中 A 的甲基化,保证 DNA 免受限制性核酸内切酶 Mbo I 的切断,同时在 DNA 复制时也起一定的辅助作用。dam 基因的变异导致腺嘌呤(A)甲基化酶活性的缺失,使腺嘌呤 (A) 不被甲基化,易于获得非甲基化质粒。 2 dcm(DNA cytosine methylase) 3 mcrA(Modified cytosine restriction protein a) 功能:mcrA 基因表达大肠杆菌防御体系中起重要作用的 mcrA 酶,这种酶能特异性地作用于外来 DNA 上的被甲基化的胞嘧啶序列,即 C5mCGG 特异序列,使之分解,对大肠杆菌本身起保护作用。mcrA 基因的变异,导致上述功能缺失,对外来 DNA 中被甲基化的胞嘧啶特异序列(C5mCGG)失去作用,有利于限制酶及甲基化酶的克隆体的稳定。 4 mcrB, C(Methyl cytosine-specific restriction) 5 mrr(Methylation requiring restriction) 功能:mrr 基因是大肠杆菌细胞防御系统中重要的基因之一,它能严格限制被甲基化的外源DNA 的介入。另外,它对限制酶 Acc I,CviR I,Hinf I (Hha II),Nla II,Pst I 以及 N6-腺嘌呤甲基化酶和 C5-胞嘧啶甲基化酶活性有明显的抑制作用。mrr 欠损株(基因型)可用于含有 N6-mA 和 C5-mC 的 DNA 的转化。另外,含有此基因型的菌株也可用于限制酶和甲基化酶的克隆体。 6 hsdM (Host specificitive defective) Map position: 99 min 功能:hsdM 基因所表达的 DNA 甲基化酶是 I 型限制酶复合体(具有对 DNA 切断和修补的双重功能)的一部分,它能使 DNA 双链上的 AA (双腺嘌呤) 甲基化,保护宿主 DNA 不被分解。hsdM 的变异使细胞内的 DNA 不被甲基化,易于获得非甲基化质粒。
03 点突变相关的基因型 1 mutS (Mutator) 2 mutT(Mutator) 功能:野生大肠杆菌在进行 DNA 复制时,细胞中的 8-OXO-dGTP 插入模板 DNA 中的 DA位点的效率几乎与插入 DC 位点的效率相同,导致 A-T 转换成 G-C,使 DNA 产生变异。而mutT 蛋白就是特异性地降解 8-OXO-dGTP 成为单磷酸盐(8-OXO-dGMP),这种单磷酸盐状态的 G (鸟嘌呤) 不能作为底物进行 DNA 合成,从而防止了上述的基因突变。 mutT 基因的变异使细胞中 8-OXO-dGTP 浓度增高,A→C 的突变几率增大,有利于利用点突变进行基因改造。 3 dut (dUTPase) 功能:dut 基因表达脱氧尿嘧啶三磷酸核苷酸水解酶(dUTPase),它能水解 dUTP 成为dUMP,使细胞体内 dUTP 的浓度维持在较低的水平,尿嘧啶(U)就不易掺入到 DNA 中,避免了基因发生 A→U 的突变。 dut 基因发生突变使 dUTPase 活性缺失,导致 dUTP 浓度升高,碱基 U(尿嘧啶)极易掺入到 DNA 中,使其发生 A→U 的基因突变,有利于利用点突变进行基因改造。 4 ung (Uracil DNA glycosylase) 5 uvrB (Ultraviolet) 功能:uvrB 基因表达核酸外切酶中的 b 亚基,这种核酸外切酶具有 DNA 的切补功能,对紫外线损伤的 DNA 有修补作用。 uvrB 基因的变异使细胞中核酸外切酶切除变异碱基的活性缺失,有利于点突变。 04 核酸内切酶相关的基因型 1 hsdR(Host specificity defective) 功能:hsdR 基因表达 I 型限制酶 EcoK (K12 株) 或 EcoB (B 株),在大肠杆菌细胞中起到 一种“抗体”的作用,对外来的各种 DNA 有严格的限制。 HsdR基因的变异导致菌株细胞 内的 I 型限制酶 EcoK 或 EcoB 活性缺失,这对于外来基因的导入及质粒转化是有利的。 2 hsdS (Host specificitive defective) 3 endA (Endonuclease) 功能:endA 基因表达非特异性核酸内切酶 I,它能使所有 DNA 双链解开,在 DNA 的复制和重组中起重要作用。 endA 基因的变异将使插入的外源 DNA 更加稳定,提取的质粒纯度 更高。 05 停止密码子相关的基因型 1 supE (Suppressor) 功能:supE 基因表达的阻遏蛋白与停止密码子 UAG 结合,使蛋白质合成停止。supE 基因 发生变异时,不能表达正常的阻遏蛋白,即使遇到停止密码子 UAG,蛋白质合成仍能继续,并使 UAG 作为一个密码子来编码 谷氨酰胺(Glutamine),从而使发生了琥珀突变(AAG→UAG)的基因对蛋白质表达得以延续,因此称 supE 为琥珀突变抑制因子。 2 supF (Suppressor) 功能:supF 基因表达的阻遏蛋白与停止密码子 UAG 结合,使蛋白质合成停止。supF 基因变异时,不能表达正常的阻遏蛋白,即使遇到停止密码子 UAG,蛋白质合成仍能继续,并使 UAG 作为一个密码子编码 酪氨酸(Tyrosine)。 06 抗药性相关的基因型
1 gyrA(Gyrase) 功能:gyrA 基因表达 DNA 促旋酶 A 亚基。DNA 促旋酶在 DNA 复制时具有使 DNA 解旋和回旋的作用。萘啶酮酸 (Nalidixic acid)、4-喹啉(4-Quinoline)等抗生素抑制 DNA 促旋酶的活性是通过与促旋酶复合体 (A2B2)中的 A 亚基的结合实现的。 gyrA 基因的变异使 DNA促旋酶 A 亚基不能正常表达,萘啶酮酸和荧光喹啉等失去结合目标,从而使该基因型的菌株具有了对萘啶酮酸(Nalr)和荧光喹啉的抗性。 2 rpsL(Ribosomal protein small subunit) 功能:细胞中的核糖体是蛋白质生物合成的场所,大肠杆菌细胞中的核糖体包含两个亚基,即 50S 亚基(23SrRNA、5SrRNA、34 种蛋白质)和 30S 亚基 [16SrRNA、21 种蛋白质(S1~S21)]。rpsL 基因就是表达核糖体 30S 亚基中的 S12 蛋白质,S12 蛋白作用于翻译的开始阶段。链霉素(Streptomycin)等抗生素的作用位点就是核糖体 30S 亚基上的 S12蛋白质,正常情况下链霉素与 S12 蛋白结合使蛋白质的生物合成不能进行,细胞停止生长。 rpsL 基因的变异使链霉素失去结合位点,从而使该基因型的菌株具有了对链霉素的抗性(Str r)。 3 Tn5(Transposon) 功能:在原核生物和真核生物基因组中都存在有可移动的 DNA 序列,一般称这段序列为转座子或转位基因,转座子上通常带有抗药性基因。Tn5 是载有卡那霉素(Kanamycine)抗性基因的转座子,当Tn5转位到大肠杆菌基因组时,能使此菌株获得卡那霉素的抗性(Kmr)。 4 Tn10(Transposon) 功能: Tn10是载有四环素(tetracycline)抗性基因的转座子。当 Tn10 转位至大肠杆菌基因组时,能使此菌株获得四环素的抗性 (Tet r)。 07 能量代谢相关的基因型 1 lacZ(Lactose) 功能:lacZ 基因是大肠杆菌中 lac 操纵子的结构基因,表达β-半乳糖苷酶,分解乳糖为半乳糖苷。β-半乳糖苷酶是由四个相同的亚基组成的,每个亚基又包含两个片断, 即α片断和ω片断,只有这两种片断同时存在时,β-半乳糖苷酶才表现出活性。lacZ 基因的变异或缺失将直接导致β-半乳糖苷酶活性缺失,细胞在只有乳糖作为碳源的培养基中不能生长,由此可以进行菌株的筛选和纯化。 2 lacZ M15(Lactose) 功能: lacZM15 是表达β-半乳糖苷酶α片断的一段基因,当 M15 缺失(△M15)时,lacZ基因虽然能表达ω片断,但不能表达α片断,β-半乳糖苷酶没有活性。当带有 lacZ(α片断)基因的 lac 操纵子通过载体 DNA(如 pUC19 DNA)转化到 lacZ△M15 基因型的细胞 (如E.coli JM109)时,在有 IPTG (异丙基-β-D-1-硫代半乳糖苷) 存在的情况下, β-半乳糖苷酶表现出活性,它能分解 X-gal (半乳糖类似物),使其呈现蓝色。因此可以通过平板上的蓝白菌落进行克隆体的鉴定。 3 lacI q (Lactose) 功能:lacI 是大肠杆菌中 lac 操纵子(Operon)的调节基因,它所表达的阻遏蛋白是 lac 操纵基因(Operator)的抑制因子,这种阻遏蛋白能与过量的乳糖结合而失去对操纵基因的抑制,使 lac 操纵子上的结构基因 lacZ(β-半乳糖苷酶)、lacY(透性酶)、lacA(乙酰基转移梅)得以正常表达。IPTG(异丙基-β-D-1-硫代半乳糖苷) 作为乳糖的类似物与 lacI 阻遏蛋白结合而使操纵基因不被抑制,因此 IPTG 经常作为 lac 操纵子的诱导剂而使用。 基因型 lacI q 是 lacI 基因发生变异而使其大量(quantity)的表达阻遏蛋白,从而使 lac 操纵基因几乎完全被抑制。利用这种基因型的菌株进行基因表达时,可以使目的基因的表达得到更有效的人为控制。 4 ara(Arabinose) 功能:ara 基因表达阿拉伯糖代谢所需的各种酶,包括:araA 表达阿拉伯糖异构酶、araB表达核酮糖激酶、araC 表达阻遏蛋白(起调节作用),araD 表达 L-核酮糖-4-磷酸差向异构酶、araE 表达低亲和型 L-阿拉伯糖转运蛋白、araF 表达 L-阿拉伯糖结合蛋白、araG 表达高亲和性的 L-阿拉伯糖转运蛋白。ara 基因的变异,使细胞不能利用阿拉伯糖进行能量代谢,可以利用此特性进行菌株筛选。 5 mtl(Mannitol) 功能:mtl 基因包括 mtlA、mtlC、mtlD 三种基因。mtlA 表达磷酸转移酶、mtlC 表达阻遏蛋白(起调节作用)、mtlD 表达甘露醇-1-磷酸脱氢酶。 mtl 基因的变异使甘露醇代谢不能进行,细胞在以甘露醇作为唯一碳源的培养基中不能生长。 6 xyl(Xylose) 功能:xyl 基因包含 xylA、xylB、xylR 三种基因。xylA 表达 D-木糖异构酶、xylB 表达木酮糖激酶、xylR 作为调节基因表达阻遏蛋白。 xyl 基因的变异使细胞不能以木糖作为碳源进行能 量代谢。 7 gal(Galactose) 功能:gal 基因表达半乳糖代谢所需的各种酶类及调节蛋白,包括:galE(17 min)表达尿苷二磷酸(UDG)半乳糖-4-差向异构酶、galK(17 min)表达半乳糖激酶、galP(64 min) 表达半乳糖透性酶、galR(62 min)表达半乳糖操纵子的阻遏蛋白、galT(17 min)表达半乳糖-1-磷酸尿苷酰转移酶、galU(27 min)表达葡萄糖-1-磷酸尿苷酰转移酶。 大肠杆菌 K12 株中通常出现的基因型是 galK 和 galT,由于这两种基因的变异使细胞不能直接利用半乳糖作为碳源。因此通过在最小培养基中添加半乳糖与否,进行菌株筛选和基因型确认。 8 srl(Sorbitol) 功能:srl 基因包含 srlA、srlC、srlD、srlR 等基因。srlA 表达磷酸转移系统相关的酶(葡萄糖醇-山梨醇透性酶、磷酸转移酶 II 等)、srlD 表达山梨醇-6-磷酸-2-脱氢酶、srlC、R 都是调 控基因,表达葡萄糖醇操纵子的阻遏蛋白。 Srl基因的变异使细胞对山梨醇的吸收和利用受到阻害,在以山梨醇作为唯一碳源的培养基中,此基因型的菌株不能生长。 08 氨基酸代谢相关的基因型 ①gpt(Guanine-hypoxanthine phosphoribosyl transferase) 功能: gpt 基因表达鸟嘌呤-次黄嘌呤磷酸核糖转移酶,参与鸟嘌呤代谢。gpt 基因的变异使鸟嘌呤不能生物合成,对菌株筛选有利。 2 thyA (Thymine) 功能: thyA基因表达胸苷酸合成酶,参与胸腺嘧啶的代谢。thyA 基因的变异可以利用胸腺嘧啶进行菌株筛选。 3 asd (Aspartate-semialdehyde dehydrogenase) 功能: asd基因表达天门冬氨酸半醛脱氢酶,催化如下反应:L 天门冬氨酸-4-半醛 + 磷酸盐 + NADP+ = L-4-磷酸天门冬氨酸 +NADPH,此反应是氨基酸共同合成路径的第二步。asd 基因的变异使天门冬氨酸合成受阻,用最小培养基进行细胞培养时,需特别添加天门冬氨酸。 4 leuB (Leucine) 功能: leuB基因表达 3(β)-异丙基苹果酸脱氢酶,作用于亮氨酸生物合成的第二步,催化反应如下:3-羧基-2-羟基-4-甲基戊烯 + NAD+→3-羧基-4-甲基-2-氧戊烯 + NADH。leuB 基因的变异导致亮氨酸生物合成受阻,在用最小培养基进行细胞培养时,需特别添加亮氨酸。 5 proA (Proline) 功能: proA基因表达γ-谷氨酸磷酸还原酶,作用于脯氨酸生物合成的第二步,反应如下: L-谷氨酸-5-半醛 + 磷酸 + NADP+→L-γ-谷氨酸-5-磷酸盐 + NADPH 。proA 基因的变异或缺失,使脯氨酸的生物合成受阻,在用最小培养基培养细胞时需要特别添加脯氨酸。 6 proB (Proline) 功能: proB 基因表达谷氨酸岩-5-磷酸激酶,它能催化三磷酸盐与谷氨酸盐结合形成谷氨酸-5-磷酸盐,是脯氨酸合成的第一步,反应如下:ATP +L-谷氨酸盐→ADP + L-谷氨酸-5-磷酸。proB 基因的变异或缺失,使脯氨酸合成受阻,在用最小培养基培养时需特别添加脯氨酸。 7 trpR(Tryptophan) 功能: trpR基因表达“trp 操纵子”的阻遏蛋白,但这种阻遏蛋白不能单独与操纵子上的操纵基因结合,只有在 L-色氨酸存在的情况下,首先与 L-色氨酸结合成复合体,然后这个复合体才能与操纵基因相结合,对 trp 操纵子起抑制作用。吲哚丙酸盐(IPA)作为 L-色氨酸的类似物也能与这种阻遏蛋白结合,但其形成的复合体没有活性,不能与操纵基因结合,因此可以把吲哚丙酸盐(IPA)作为 trp 操纵子表达的诱导剂。trpR 基因的变异,使 trp 操纵子的阻遏蛋白不能表达,有利于 trp 操纵子的蛋白表达。 8 lys (Lysine) 功能: lys基因分布于大肠杆菌基因组图的 17 ~ 191 min 的 5 个位置上,包括 lysA (61 min)、lysC (91 min)、lysP (46 min)、lysT (17 min)、lysX (60 min),它们的功能如下:lysA 表达二氨基庚二酸脱羧酶、lysC 表达天冬氨酸激酶、lysP 是调节赖氨酸转运的基因、转录赖氨酸 tRNA、lysX 负责赖氨酸排泄。lys 基因的变异使赖氨酸的生物合成不能进行,用最小培养基培养时需额外添加赖氨酸 。 9 metB(Methionine) 功能: metB基因表达胱硫醚γ-合成酶,催化反应如下:0-琥珀酰-L-高丝氨酸 + L-半胱氨酸→胱硫醚 + 琥珀酸盐,是甲硫氨酸生物合成的第二步。metB 基因的变异使甲硫氨酸的生物合成受阻,用最小培养基培养时需特别添加甲硫氨酸。 10 cysB (Cysteine) 功能: cysB基因表达一种阻遏蛋白,对半胱氨酸生物合成所需的各种酶的表达起调节作用。cysB 基因的变异有利于半胱氨酸的生物合成。 11 thr(Thronine) 功能: thr包含三种基因,即 thrA、thrB 和 thrC。thrA 表达天冬氨酸激酶及 I-高丝氨酸脱氢酶,thrB 表达高丝氨酸激酶,thrC 表述苏氨酸合成酶。thr 基因的变异使细胞不能合成苏氨酸,用最小培养基培养时需添加苏氨酸。 09 维生素代谢相关的基因型 1 bioH(Biotin) 功能:bioH 基因所表达的蛋白有两种功能:①催化前生物素到生物素的转化;②对庚二酰CoA(辅酶 A)有优先的阻害作用。 bioH 基因的变异使细胞不能自身合成生物素,在最小培养基中必须添加生物素,细胞才能正常生长。 2 thi(Thiamin) 功能:thi 基因包含有 thiA、thiB、thiC、thiD、thiK、thiL 等。thiA 表达硫氨素噻唑转运蛋白、thiB 表达硫氨素磷酸盐焦磷酸化酶、thiC 表达硫氨素嘧啶转运蛋白、thiD 表达磷酸甲基化嘧啶激酶、thiK 表达硫氨素激酶、thiL 表达硫氨素甲磷酸激酶。 thi的变异使硫氨素的生物合成不能进行,最小培养基中必须添加硫氨素(VB1),细胞才能正常生长。 10 蛋白酶相关的基因型 1 lon(long form) 功能:lon 基因表达 ATP 依赖型蛋白分解酶(La),它对外源的异型蛋白质具有特异性的分解作用。lon 基因的变异或缺失,使细胞中的这种异型蛋白质分解酶不能得到表达,这对于保持克隆体目的蛋白的稳定是非常有利的。 2 ompT(Outer membrane protein) 功能:ompT 基因表达特异性的外膜蛋白分解酶,它特异性地分解与细胞膜结合的含铁肠菌素受体蛋白。ompT 基因的变异使膜结合性蛋白分解酶活性缺失,有利于融合蛋白的表达。 11 物质转运结合相关的基因型 1 tonA(Transport)=fhuA(Ferric hydroxamateuptake) 功能: tonA 和 fhuA 基因处于基因组图同一位点,它们的作用也相同,都是表达外膜受体蛋白。这种受体蛋白与铁络合物结合,并与 tonB 蛋白相互协调作用,把铁化合物运至细胞质中。另外 tonA、B 受体蛋白还能与大肠杆菌素 M、噬菌体 T1、T5、φ80 等进行不可逆结合,而使细胞具有一定的抗菌作用。tonA 和 fhuA 基因的变异使细胞对铁离子的吸收受到阻害,同时使细胞对某些抗菌素及噬菌体更敏感,有利于质粒转化和菌体筛选。 2 tsx(T-six) 功能: tsx 基因表达 T6 噬菌体和大肠杆菌素 K 的受体蛋白,它结合于细胞膜的外表面,对T6 噬菌体和大肠杆菌素 K 进入细胞起关键作用。另外 tsx 受体蛋白还有与核苷酸特异性结合的功能,是核苷酸特异性运输通道的第一步,在核苷酸的吸收方面也起重要作用。tsx 基因的变异使外界某些噬菌体及大肠杆菌素等对细胞的侵噬变得困难,有利于细胞基因组的稳定。 3 cysA (Cysteine) 功能: cysA 基因表达硫酸盐/硫代硫酸盐转移蛋白,参与细胞对硫酸盐的吸收与转运,通过cysA 蛋白转运的硫酸盐将参与半胱氨酸的生物合成。cysA 基因的变异使半胱氨酸的生物合成受到影响,在培养此基因型的菌株时要注意添加半胱氨酸。 12 其他 1 deoR (Deoxyribose) 功能: deoR是大肠杆菌中 deo 操纵子的调节基因,它表达的阻遏蛋白对 deo 操纵子具有重要的调控作用。deo 操纵子位于大肠杆菌基因图谱 100 min 的位置,它含有 deoA、 deoB、 deoC和 deoD等结构基因,分别表达 DNA 代谢所需的酶类,即胸腺嘧啶磷酸化酶、磷酸转位酶、脱氧核糖磷酸醛缩酶和嘌呤核苷磷酸化酶。deoR 基因的变异,使 deo 操纵子的阻遏蛋白不能表达,宿主细胞合成大量的 dCTP,可以选择性地改善大分子 DNA 的转化。 2 traD(Transmissibility) 功能: traD基因不属于大肠杆菌基因组 DNA 范围,它是存在于 F 因子上的一段基因。 traD基因在大肠杆菌的结合及 F 因子的传递方面发挥作用。 traD基因的变异使大肠杆菌细胞虽然能够结合,但 F 因子不能在细胞间发生转移,从而保证了宿主细胞和导入质粒的稳定性。 3 hflC(High frequence of lysogenization) 功能: hflC基因表达一种高效溶原蛋白,它能使λ-噬菌体侵入大肠杆菌细胞后与基因组DNA发生溶原反应,导致噬菌体 DNA 插入到细胞基因组 DNA 中。 hflC基因的变异能避免上述的溶原反应,可以保持宿主基因组及插入质粒的稳定。 4 minA、B (Minicell) 功能: minA、B 基因是促进微细胞 (不含 DNA) 形成的相关基因。 minA、B基因的变异阻害了微细胞的形成,可以提高克隆体的表达效率。 5 relA(Relaxed) 功能: relA是松弛调节基因,对 RNA 的合成具有调节抑制作用。同时 relA 基因还表达 ATP: GTP3'-焦磷酸转移酶,负责在氨基酸饥饿状态下鸟苷 3',5'-二磷酸 (ppGpp) 的合成,以适应饥饿环境。 relA 基因的变异对目的基因的转录有利。 6 glnV(Glutamine) 功能: glnV基因专门负责转录谷氨酸 tRNA(转运 RNA), glnV基因的变异使以谷氨酸为主的蛋白质的合成受到阻害。 7 tyrT(Tyrosine) 功能: tyrT基因专门负责转录酪氨酸 tRNA(转运 RNA), tyrT基因的变异使以酪氨酸为主的蛋白质的合成受阻。 使用时注意: 基因工程中,经常使用的大肠杆菌几乎都来自于 K-12 菌株,最近也经常使用由 B 株及 C株来源的大肠杆菌。 ● 大肠杆菌 B 株原来就为 lon-,另外 MV1184 株不具有琥珀抑制基因 (Amber supperssor free),由于这些都是原始菌株所不具备的基因,因此不在基因型中加以表示,要注意。返回搜狐,查看更多 |



【本文地址】
今日新闻 |
推荐新闻 |