顶刊综述 |
您所在的位置:网站首页 › windowsprojectedfilesystem作用 › 顶刊综述 |
顶刊综述
|
导读 大脑和胃肠道是负责感应、传递、整合和回应来自内外环境信号的感觉器官,由于具有这种功能,大脑和肠道内的免疫细胞会持续感应环境因素,从而引发能反映身体生理状态的反应。最近有研究表明,沿肠道-脑轴的交互信号可调节炎症性伤害感受、炎症反应和免疫稳态。在本综述中,我们将讨论沿肠道-脑轴传递的炎症信号的分子和细胞机制,从而进一步强调肠道和大脑在炎症相关性疾病中的相互作用。 论文ID 原名:Signaling inflammation across the gut-brain axis 译名:通过肠-脑轴传递的炎症信号机制 期刊:Science IF:63.714 发表时间:2021.11 通讯作者:Gulistan Agirman 通讯作者单位:加利福尼亚大学洛杉矶分校 DOI号:10.1126/science.abi6087 综述目录 1. 前言 2. 肠-脑轴中交互的炎症信号 3. 通过肠-脑轴传递的炎症信号 3.1 系统体液途径 3.2 细胞免疫途径 3.3 神经途径 4. 炎症相关性疾病中的肠-脑轴交流 5. 总结和未来展望 主要内容 1. 前言 胃肠道(GI)和中枢神经系统(CNS)可持续感应内外环境变化,因此,GI和CNS拥有复杂的细胞网络,包括免疫细胞和神经元细胞,以便监测有害刺激和协调局部和全身的炎症反应。炎症信号在传入(“肠-脑”)和传出(“脑-轴”)方向上都发生在肠-脑轴上,以此传递宿主的健康状况,并激发有利于恢复稳态或放大炎症反应的调解反应。 本综述讨论了在稳态状况和炎症性相关疾病中炎症信号通过肠-脑轴被感知和传递的分子和细胞机制。 2. 肠-脑轴中交互的炎症信号 GI常年暴露于食物和环境的刺激因素中,同时还受内源性肠道微生物群和它们的分子产物所影响。在GI中,有3种物理屏障协同作用以阻止管腔内容物失控地进入肠道固有层和血液中,包括由杯状细胞产生的厚黏膜层、半透性单层肠细胞层和由内皮细胞、周细胞和肠胶质细胞组成的肠血管屏障(GVB)。此外,肠道是体内最大的免疫细胞群,它们可高效地感应肠道环境中的刺激因素(表1)。 先天性和适应性免疫系统通过协同工作来应对肠道的应激反应,先天性免疫细胞包括树突状细胞(DCs)、M细胞、巨噬细胞、肥大细胞、自然杀伤细胞,以及1型、2型和3型先天性淋巴细胞(ILCs),这些细胞作为肠道应激反应的第一道防线。在胃肠道淋巴样结构中,适应性免疫细胞在DC依赖性抗原呈递时可分化为各种效应亚型。细胞毒性CD8+T细胞和CD4效应细胞[辅助型T1细胞(TH1细胞)、TH2细胞和TH17细胞]和调节性T细胞(Treg细胞)不仅在局部起作用,还可以迁移到其他组织,包括大脑。 肠神经系统中的神经元和胶质细胞均参与肠道免疫,在支持肠神经细胞的同时,肠胶质细胞通过分泌RET受体配体来保护肠道屏障,该配体可触发IL3依赖性的白细胞介素-22 (IL-22) 的产生。肌层黏膜内的组织保护性巨噬细胞通过BMP2-BMPR信号与肠神经元密切互动,以此促进神经元存活,同时限制炎症诱导的肠蠕动功能障碍。 然而,神经元-免疫反应并不总是有利的,例如,若黄病毒感染了细胞毒性CD8+T细胞,会引起肠神经元细胞死亡和慢性肠功能障碍。感染之后,非炎症性刺激会加剧肠运动障碍,导致肠神经元永久性损伤和过敏。鉴于肠道神经元是外源性神经纤维的目标,它们的功能紊乱可以改变肠-脑互通和CNS驱动的肠道控制。 在稳态条件下,CNS因血脑屏障(BBB)与外周隔离,BBB与GVB具有分子和细胞相似性,其局部免疫功能主要由胶质细胞支撑(表1)。小胶质细胞是CNS中主要的常驻免疫细胞,通过在脑实质中表达而较全面地感受大脑信号,该细胞具有抗原表达能力,并与浸润的淋巴细胞结合以限制病原体的侵袭。除了细胞因子,小胶质细胞还感应神经调质和递质,从而引发免疫反应,这取决于特定的配体-受体相互作用和炎症发生的特定位置。像ILC2一样,小胶质细胞可表达高水平的β2AR,通过去甲肾上腺素激活后其运动方式和形态会改变,尽管这些功能变化结局尚不清楚。 星形胶质细胞是CNS中数量最多的胶质细胞,具有促进血脑屏障完整性和参与免疫调节的重要功能。 在神经炎症的背景下,星形胶质细胞释放转化生长因子-β(TGFβ),它能改变小胶质细胞中的染色质景观,以限制它们的激活。值得注意的是,星形胶质细胞来源的TGFβ和IL-33可刺激小胶质细胞的突触剪切活动,这表明神经炎症反应可以导致神经元突触的异常消除,并改变大脑回路。另一方面,激活的小胶质细胞可通过分泌IL-1α和肿瘤坏死因子(TNF)而诱导星形胶质细胞中的神经毒性A1状态。 除了脑内的免疫细胞,大量白细胞(包括DCs、巨噬细胞、ILC2、B细胞和T细胞)也存在与CNS和外周之间的脑膜层中(表1)。虽然通常报道脑膜免疫细胞亚型是在感染或损伤时被激活,但其核心功能才刚刚被阐明。脑膜T细胞通过分泌IL-4和IFN-γ来支撑CNS的认知功能,星形胶质细胞与脑膜T细胞互通,通过分泌IL-12和IL-23来增强TH1和TH17的反应,或通过分泌IL-27来限制TH17的细胞活性和促进产生IL-10的T细胞。这些CNS相关的免疫细胞大部分来源于外周,尽管脑膜单核细胞、中性粒细胞和B细胞的亚群直接来自于颅骨和脊椎的骨髓中。 大脑中局部产生的免疫细胞与血液来源的免疫细胞分子类型不同,并与炎症相关的转录谱降低相关,这表明它们可能在CNS损伤时负责调节神经炎症反应。此外,还有一些边界相关的巨噬细胞,如血管周围和脉络膜丛巨噬细胞,但它们的免疫功能和分泌细胞仍不清楚。总之,这些免疫细胞和非免疫细胞形成了复杂的相互作用网络,以调节CNS和IG的炎症反应。 表1 肠道和CNS驻留的免疫细胞参与肠-脑轴的炎症信号传导。
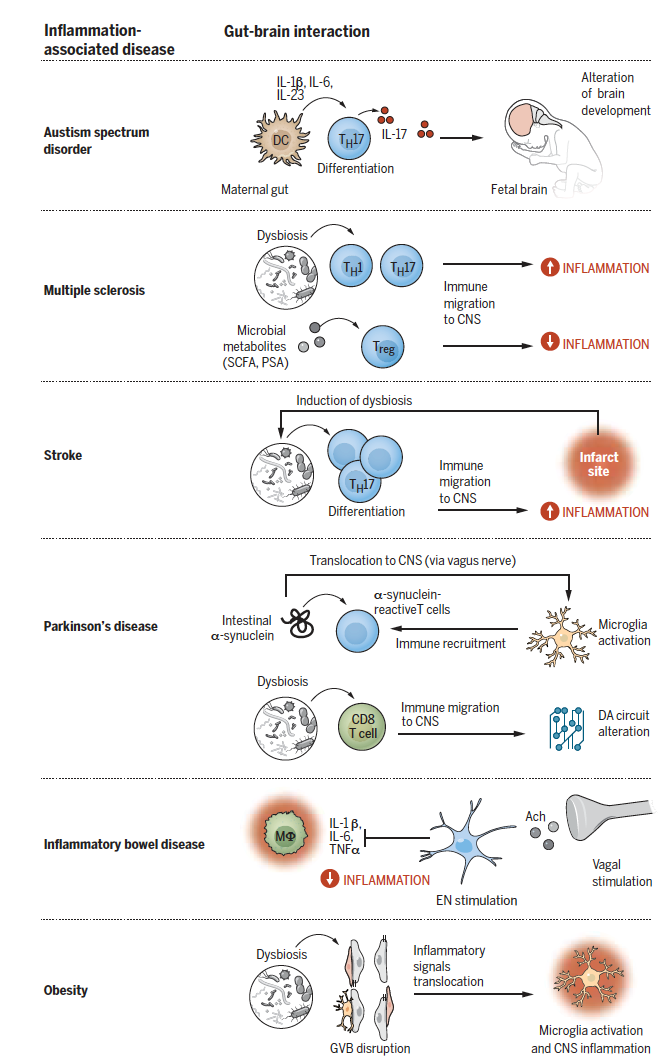
对肠道和CNS损伤的反应是由肠道和大脑免疫细胞网络之间的相互作用介导的,Ach,乙酰胆碱;CNS,中枢神经系统;ILC,先天性淋巴细胞;NK,自然杀伤细胞;SCFA,短链脂肪酸;TH,辅助型T细胞;Treg,调节性T细胞。 3. 通过肠-脑轴传递的炎症信号 肠-脑轴包括双向和连续的交互反应来调节分子、细胞和功能状态,炎症信号在IG和CNS之间可以通过三条平行但相互连接的通信通途来传递信号(图1)。 图1 沿肠-脑轴的炎症信号通路。 炎症信号在肠和CNS之间通过三个主要途径进行双向传递:(顶端)体液途径,肠源性炎症因子的全身释放可以改变BBB的完整性,导致大脑发育缺陷,此外,炎症诱导的HPA轴活动可触发全身释放糖皮质激素,从而改变肠道功能。(中层)细胞免疫途径,激活的肠免疫细胞可转运到CNS中而促进或抑制神经炎症反应,对CNS损伤的应激反应可以改变肠道微生物群和刺激炎症免疫细胞产生,这些细胞随后又迁移到CNS而加重神经炎症反应,来自肠道菌群的代谢物可调控脑驻留免疫细胞的发育和神经炎症反应。(底端)神经途径,炎症敏感传入迷走神经和DRG神经元的激活可引发HPA激活、疾病行为和内脏痛觉有关的中枢神经回路,传出神经元向免疫细胞发出信号后,通过抑制促炎巨噬细胞、调节M细胞的功能以限制病原体入侵,并诱导保护性微生物的发展等方式来抑制肠道炎症。Ach,乙酰胆碱;ACTH,促肾上腺皮质激素;BBB,血脑屏障;CRH,促肾上腺素皮质激素释放激素;DMV,迷走神经背侧运动核;DRG,背根神经节;EN,肠源性神经元;GV,肠-血管屏障;HPA,下丘脑-肾上腺-垂体轴;MΦ,巨噬细胞;NG,节状神经节;NK,自然杀伤细胞;NTS,孤束核;PC,浆细胞;SC,脊髓;SFB,分节丝状细菌;TH,辅助型T细胞;Treg,调节性T细胞;VN,迷走神经。 3.1 系统体液途径 由局部感染、功能失调或食物抗原引起的肠道炎症会诱导促炎细胞因子的释放,如IFN-γ、IL-1β、IL-6和TNFα,以及其它免疫因子,若不加以控制,这些因子会产生严重的肠外结局。循环中的促炎因子可通过破坏紧密连接的上皮而损害GVB和BBB等屏障,致使肠通透性增加、或“肠漏”,辅之以CNS屏障的破坏,为肠源性分子、毒素和病原体到达脑实质,激活局部免疫细胞,并引发神经炎症提供了通道。例如,细菌壁组分脂多糖(LPS)的全身水平升高与小胶质细胞激活、神经元细胞死亡、认知障碍和细胞因子介导的疾病行为(运动能力降低、食物摄入量减少和社会退缩)相关。 一些神经系统疾病与肠道通透性增加有关,微生物依赖性肠道屏障功能恢复可以改善小鼠的选择性行为异常。除了局部影响肠道通透性,肠道微生物及其产物还可以影响CNS的神经免疫调节功能。小胶质细胞、星形胶质细胞和边界相关性巨噬细胞的发育、成熟和功能均受到微生物依赖信号的调节,缺乏免疫启动则会导致对脑损伤和炎症刺激的反应缺失。例如,短链脂肪酸(SCFAs)可调节BBB通透性和小胶质细胞功能,并最终调节宿主的神经炎症反应。 SCFAs还诱导肠内分泌细胞 (EECs) 产生和释放GLP1和PYY激素,这些激素可间接调节CNS对餐后饱腹感和情绪状态的影响,这两者在炎症相关的糖尿病和肥胖中都存在失调。来自肠道的色氨酸代谢物构成了另一个调节宿主炎症反应的微生物群落依赖信号的例子,色氨酸代谢产物可靶向作用于芳香烃受体,以此局部增强肠道屏障,并通过诱导星形胶质细胞IFN-I信号传导而减轻CNS中的炎症。大多数肠道或微生物源性因子尚未在CNS功能的背景下进行研究,因为人们认为它们既不跨越GVB,也不跨越BBB,然而,这些屏障的破坏可能会使一些因子进入CNS,并与其免疫细胞和神经元产生作用。需要进一步的研究来评估肠源性分子和CNS细胞类型之间的特异性相互作用,尤其是在炎症状态下的作用。 到目前为止,下丘脑-垂体-肾上腺(HPA)轴的活动是肠-脑轴的主要传出体液信号轴。环境应激源或肠道炎症的外周反应整合到CNS中触发HPA轴,以此协调肾上腺糖皮质激素的释放。这种有效的应激诱导激素可以通过调节肠道免疫细胞活性、肠道功能和微生物组成来恢复体内平衡或促进GI功能紊乱。 应激诱导的失调可反过来通过TH17细胞依赖性释放IL-17A而引发肠道炎症,从而促进应激反应的前馈激活。肠道微生物群也参与了稳态过程中对HPA轴的调节,因为微生物群缺乏会加剧HPA轴对中等压力的反应活性。引流脑脊液的脑膜淋巴管可能是分子从CNS进入外周刺激或启动外周器官(包括GI)免疫反应的另一途径,然而,这种通路在肠-脑交流中的作用仍然是假定的,还需进一步研究证实。 3.2 细胞免疫途径 免疫因子除了通过肠-脑轴的内分泌途径传导信号外,肠道免疫细胞本身也能直接调节神经免疫稳态和大脑对炎症的反应。肠道抗原刺激B细胞分化为分泌免疫球蛋白A(IgA)浆细胞,以控制肠道微生物群落的功能。 神经系统自身免疫性疾病可诱导肠道IgA+浆细胞大量迁移到大脑和脊髓,以IL-10依赖性方式减轻神经炎症。尽管招募细胞的信号分子尚待确定,但它们在大脑中的存在已在稳态中被证实,它们定位于硬脑膜静脉窦附近,以保护CNS免受周围病原体的侵入。通过利用肠道抗原促进IgA+ B细胞产生和神经免疫抑制,这些发现可能可为治疗神经免疫紊乱开辟新的途径。 CNS的局部免疫细胞也可由肠源性细胞编程,例如,来自肠道可产生IFN-γ的脑膜NK细胞子集,就可促进表达LAMP1和TRAIL的神经免疫调节性星形胶质细胞的发育,此外,这些肠道来源的脑膜NK细胞的功能及其调控星形胶质细胞发育的能力受到肠道微生物群的调节。与在稳态时对神经免疫状态的有利影响相反,肠道免疫细胞还可直接促进CNS内的神经炎症反应。例如在多发性硬化症(MS)或帕金森病(PD)中,肠道内的免疫细胞可被宿主或微生物来源的抗原激活,并迁移到CNS中,随后加重炎症反应效果。虽然有几种免疫细胞从肠道浸润到大脑的例子,但在传出方向的细胞通信机制仍尚不十分清楚,需要进一步的研究来揭示CNS炎症在疾病中是如何导致肠道免疫细胞激活的,并确定肠道免疫细胞迁移到大脑后如何促进神经炎症发展的。 3.3 神经途径 连接CNS和GI的神经元可传递炎症信号,从而引发CNS的反应以便恢复体内平衡。肠道传入迷走神经在肠道内的感觉微环境有感知炎症调节因子的受体,迷走神经传入纤维的细胞体位于结节神经节中并投射到脑干的孤束核,然后由孤束核将信号传递给大脑的高级区域。炎症介导的迷走神经传入纤维可刺激触发HPA轴并激活与疾病行为相关的神经回路。选择性肠道病原体能够操纵炎性IL-1β信号传到迷走神经,以防止下丘脑刺激引起的厌食反应。 在这种情况下,抑制疾病行为的神经感觉途径可降低致病体毒性,从而促进其传播。肠道炎症的中央性调节则通过迷走神经背侧运动核的迷走神经下行路径发生,胆碱能迷走神经传出纤维可刺激肠神经元,抑制巨噬细胞释放炎症因子IL-1β、IL-6、IL-18和TNF-a。虽然黏膜巨噬细胞与迷走神经末梢的距离较近,但尚不清楚迷走刺激是否能直接调节肠免疫细胞或间接向肠神经元、胶质细胞或肠细胞传递信号。 痛觉是炎症的主要表现形式之一,内脏敏感性主要通过脊髓背角处的背根神经节(DRG)中的痛觉传入神经传递给CNS 。活化的肠免疫细胞可释放5-HT、组胺、IL-6和TNF-a等炎症介质,从而增强电压门控通道的激活,并诱导伤害感受器神经元的兴奋性。DRG神经元通过脊髓将感应到的有害刺激传递到大脑,并激活调节疼痛信号感知、整合和情感成分的神经回路。同时,伤害性神经元可刺激脊髓小胶质细胞、星形胶质细胞和少突胶质细胞,这可进一步调节电压通道的门控特性,以增强痛觉感受器神经元的敏感性。 此外,来自大脑的传出信号可激活脊髓背角的抑制通路,从而调节痛觉神经元的活动。 除了促进炎症性疼痛,DRG痛觉感受器神经元已被证明可以防止肠道病原菌感染。肠神经支配的TRPV1+感受性神经元能够感知沙门氏菌(Salmonella)感染,并释放CGRP来减少作为致病传入口的Peyer 's斑块中M细胞的丰度。TRPV1+痛觉感受器神经元的激活也增加了菌群中节段性丝状细菌(SFB)的丰度,从而限制病原体的定植、入侵和传播。鉴于病原体可以操纵迷走神经传入信号来预防疾病行为,这种机制可能进一步有利于病原体的宿主传播,这些效应是否需要来自CNS的调节性反馈是未知的,还需要更多的研究来探索脊髓传出通路的肠调节机制。 4. 炎症相关性疾病中的肠-脑轴交流 一些神经系统疾病伴随着肠道炎症和促炎症血清学特征,这些特征与大脑和行为受损相关,包括认知能力下降、自闭症谱系障碍(ASD)和肥胖的神经行为症状(图2)。衰老相关的认知功能下降与慢性低级别的肠道和中枢炎症状态相关,包括CNS中小胶质细胞、T细胞和边界相关巨噬细胞水平增加以及肠道微生物群的改变,衰老相关性B细胞也从外周侵入脑膜并分化为产生IgM的 浆细胞。 值得注意的是,肠道失调在CNS免疫改变之前可引起肠道通透性改变并诱发神经炎症症状,这提示肠道失调有利于炎症因子通过肠-脑轴的体液信号传导。
图2 炎症相关性疾病中的肠-脑轴交流。 肠源性炎症分子通过胎-孕界面的体液转运可导致后代ASD相关的社交障碍,肠道微生物失调可促进肠道免疫细胞的激活,进而转运到CNS,以增强MS、中风和PD等疾病的神经炎症反应。迷走神经刺激可通过抑制炎症性巨噬细胞的功能而抑制功能性肠紊乱引起的炎症反应,代谢性疾病如肥胖与GVB的破坏有关,这可能导致CNS中驻留性免疫细胞的激活。Ach,乙酰胆碱;CNS,中枢神经系统;DA,多巴胺;DC,树突细胞;EN,肠源性神经元;GVB,肠-血管屏障;MΦ,巨噬细胞;PSA,多糖A;SCFA,短链脂肪酸;TH,辅助型T细胞;Treg,调节性T细胞。 在ASD小鼠模型中,妊娠期间母体感染或免疫激活可诱导肠道中产生IL-17的TH17细胞以DC依赖性方式分化。循环性IL-17穿过胎盘屏障后可改变CNS的发育,从而影响自闭症后代的大脑和行为内表型,这项研究证明了肠道炎症因子可以通过胎-孕界面的体液传递而刺激神经发育异常的概念,因此,通过肠-脑轴神经通路的微生物依赖性信号可能有助于改善ASD模型中的神经发育异常,例如,用罗伊氏乳杆菌(Lactobacillus reuteri)治疗可以通过迷走神经元向微生物-肠道-大脑轴传递信号,从而纠正社交障碍。 饮食诱导的肥胖构成了另一种炎症病理状态,并通过肠-脑轴诱导细胞和神经行为的改变。以高饱和脂肪和高糖为特征的西方饮食可改变肠道屏障标记物的表达和降低EEC衍生型GLP1的表达,还可通过小胶质细胞释放IL-1β、IL-6和TNF-a而诱导中基底下丘脑炎症,并引发对食欲调节激素瘦素的中枢抵抗,进而促进焦虑样行为。在肠道微生物缺乏的情况下,下丘脑炎症可被抑制,而通过移植高脂饮食治疗小鼠的粪便内容物到无菌小鼠后,受体小鼠会表现出行为异常,提示饮食诱导的肠道微生物失调可能有助于CNS表型的发展。 肠道免疫细胞迁移到CNS也可能有助于神经和神经退行性疾病的发生,如MS、PD和中风(图2)。在MS中,肠道菌群促进了肠内髓鞘活性TH1细胞、TH17细胞和Treg细胞的发育,随后这些细胞会分别迁移到CNS中促进或抑制炎症发生。虽然细菌代谢物如多糖A和SCFAs可以诱导Treg细胞的产生,但尚不清楚具体的微生物成分是如何激活促炎T细胞的产生。 在啮齿类动物PD模型中,肠内突触核蛋白原纤维可引起局部炎症,而炎症状态可促进它们在肠道神经元中聚集,聚集后又可通过迷走神经从肠道传播到大脑。被激活的小胶质细胞随后招募肠内突触核蛋白特异性T细胞到黑质,在那里它们会促进进一步的神经炎症反应。肠道炎症本身也可导致CD8+ T细胞浸润,从而扰乱脑内多巴胺能通路。 卒中后肠道菌群失调诱导TH1和TH17细胞在肠道内增殖,并诱导肠源性T细胞和单核细胞向缺血性脑组织中迁移,从而进一步加重神经炎症反应。 微生物失调可能是由自主神经系统(ANS)的失调引起的,ANS失调可降低黏膜屏障的完整性,肠道缺血再灌注可导致硝酸盐过量产生,从而导致肠杆菌科(Enterobacteriaceae)水平升高。给予降低硝酸盐生成或呼吸的药物可以抑制肠杆菌水平升高,并减轻细菌脂多糖引起的全身炎症,从而达到改善脑梗死的目的。未来需要进一步的研究来确定引起MS和PD菌群失调和肠道炎症的特定刺激因素。 对肠-脑轴的理解可促进肠道性疾病治疗的发展,如炎症性肠病 (IBD) 和肠易激综合征 (IBS) 。如前所述,迷走神经通过胆碱能传出纤维抑制巨噬细胞释放炎症细胞因子而发挥抗炎作用,因此,迷走神经刺激 (VNS) 可降低大鼠IBD模型中的肠道免疫细胞和炎症因子的表达。即使在IBS中,VNS也能改善便秘和腹痛,并降低循环中的IL-6和TNF-α水平。因此,进一步了解参与肠-脑轴炎症信号的分子和细胞因子,将可能揭示能改善肠道和神经性疾病内表型的其他候选标志物和治疗方法。 总结和未来展望 炎症信号可通过肠-脑轴双向传递,对于调节炎症相关性疾病的生理行为和病理改变具有重要意义。在传入方向上,肠道免疫细胞可以被招募到大脑中,但感知最初炎症信号的方式及其在大脑中的作用机制尚不十分清楚,而大脑如何在传出方向调节肠道炎症也不清楚。除了ANS功能障碍外,通过传出迷走神经细胞发出信号和脑膜淋巴管将脑源性循环因子运输到外周,以及通过HPA轴的间接相互作用都是这种通信的潜在途径。 肠道菌群是肠-脑轴中免疫细胞的关键调节因子,其生物性失调与多种炎症相关疾病和风险因素有关,包括衰老、饮食和压力,然而,肠道菌群失调是否在这些情况下起因果作用则是一个无定论的问题,确定特定的肠道微生物抗原和代谢物如何影响肠道免疫细胞或如何通过BBB影响CNS的机制将有助于解决这个问题,此外,我们还应该考虑到肠道菌群在环境条件下的变化,因此,微生物依赖的肠-脑信号调节变化可能是不同饲养环境小鼠模型中肠道、神经免疫和行为症状严重程度差异的基础,这使得研究结果的临床转化具有一定的挑战性。解码沿肠-脑轴的炎症信号的基本机制对于我们理解神经免疫通信是至关重要的,并有助于发展GI和神经系统疾病中的免疫调节性治疗。 |


【本文地址】
今日新闻 |
推荐新闻 |