细胞信号通路详解之Wnt信号通路 |
您所在的位置:网站首页 › motr英文 › 细胞信号通路详解之Wnt信号通路 |
细胞信号通路详解之Wnt信号通路
|
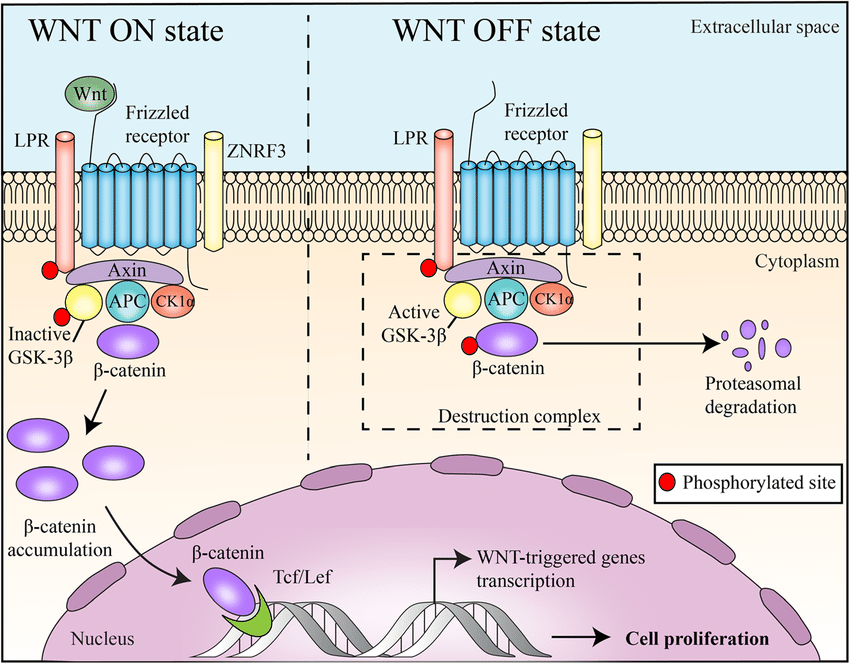
Wnt/Ctnn-Beta 通路是人们最了解的Wnt 信号通路,在进化过程中高度保守。WNT/Ctnn-Beta 途径由WNT1 、WNT3 、WNT3a 、WNT7a 和WNT8 激活,并参与转化。在没有Wnt 信号的情况下,Beta-Ctnn 与含有CK1Alpha (酪蛋白激酶-1-Alpha)、GSK3Beta (糖原合成酶激酶-3-Beta)、Axin (Axis抑制剂)和APC (腺瘤性息肉病结肠)蛋白的细胞质复合物相结合。这促进了Beta-Ctnn 的磷酸化及其与Beta-TRCP (Beta-转导蛋白重复序列蛋白)的相互作用,导致Beta-Ctnn 被泛素化,随后被蛋白酶体降解(参考文献1)。在Wnt 信号存在的情况下,Wnt 与其受体Frizzled 结合,从而导致Dsh (Dishevelled)蛋白的激活。在哺乳动物中的所有器官中,Dishevelled 蛋白家族成员都含有Dsh1 ,Dsh2 ,Dsh3 。这些家族成员有三个高度保守的结构域,包括一个N 端DIX 结构域,名字来源于Dsh 和Axin ,一个中央PDZ 结构域,是Postsynaptic Density-95 、Discs-Large 和Zonula occludens-1 的首字母缩写,以及一个C- 末端的DEP 。被激活的Dishevelled 蛋白增强了GSK3Beta 的磷酸化,从而抑制了GSK3Beta ,导致游离的未磷酸化Beta-Ctnn 在细胞质中积累,然后这些Beta-Ctnn 转运到细胞核。在WNT 信号转导前的细胞核中,LEF (Lymphoid Enhance Factor)和TCF (T-Cell factor)等与靶基因启动子/ 增强子区域的特异性序列结合,并与Groucho 和CTBP (COOH末端结合蛋白)一起,通常起到抑制基因表达的作用。WNT 信号导致核内Beta-Ctnn 水平升高,从而与TCF/Lef 结合,促进转录机制的改变,从而导致几个靶基因的激活。蛋白质从钙粘附素结合池转移到细胞质池可以增加可用于激活靶基因的游离Ctnn-Beta 数量。转录激活是通过Ctnn-Beta 与组蛋白乙酰转移酶CBP (CREB结合蛋白)、染色质重塑SWI/SNF 复合物以及与Pyg (Pygopus)和BRG1 结合的Bcl9 相互作用介导的。CHIBBY 直接与Ctnn-Beta 的C- 末端区域相互作用,通过与LEF1 竞争结合Ctnn-Beta 来抑制Ctnn-Beta 介导的转录激活(参考文献4,5和6)。
目前已有多个基因被确定为Ctnn-Beta/TCF 转录调控的靶基因。其中包括基质金属蛋白酶-7 (MMP-7)、尿激酶型纤溶酶原激活剂受体(UPAR)、CD44 、c-Myc 、c-jun 、Fos 相关抗原-1 (Fos-Related Antigen-1)、细胞周期蛋白D1 (Cyclin-D1)、过氧化物酶体增殖性激活受体- δ(PPAR-Delta)、转录因子-1 (TCF1)、纤维连接蛋白(FN)、纤连蛋白(Slug)、胃泌素(Gastin)、环氧合酶-2 (COX2)和层粘连蛋白5 (Laminin 5)的γ2 链。此外,TCF/Lef 和Ctnn-Beta 复合物可能与其他信号通路激活的因子协同作用,改变细胞重塑过程。其中许多基因,包括CCND1 和c-Myc ,在细胞生长、增殖和分化中起关键作用,并且在结肠癌中被异常激活。Ctnn-Beta 诱导的c-Myc 基因可诱导p53 表达,p53 上调p21WAF1 和p130/RB2 ,从而导致细胞生长停滞。Ctnn-Beta 的另一个靶点是Vimentin ,一种参与细胞迁移的蛋白质。在人乳腺肿瘤细胞中,Vimentin 蛋白是CtnnBeta/TCF 反式激活途径的直接靶点。WNT 信号可通过上调Caspase 抑制剂Survivin 等抗凋亡蛋白来阻止细胞凋亡,并通过上调血管内皮生长因子(VEGF)来刺激血管生成。一些能够降解细胞外基质的蛋白酶,如Matrilysin/MMP7 和MMP26 ,以及细胞粘附分子,如CD44 和NrCAM (神经元细胞粘附分子),都是WNT 的靶基因,并且可以促进肿瘤的侵袭和转移。经典WNT 信号通路还调控NRSF/REST 和ENC1 (外胚层-神经皮质(带有BTB样结构域)-1)基因,从而控制祖细胞。Cldn1 (Claudin-1)也参与了Ctnn-Beta-TCF/Lef 信号通路,Cldn1 的高表达可能在大肠癌的发生发展中起一定作用。几个WNT 信号通路的靶基因可以编码WNT 信号转导系统的组份。beta-TRCP 和泛素结合酶UbC4/5E2 的表达的增加,可能在WNT 信号转导中起负反馈作用,这两个蛋白均参与Ctnn-Beta 降解。最近的研究发现Wnt 信号通路的抑制剂有四个家族:FRP (Frizzled相关蛋白)、Cer (Cerberus)、WIF1 (Wnt抑制因子-1)和Dkk1 (Dickkopf-1)。Cerberus 和WIF1 可直接与WNT 相互作用并抑制WNT 。FRP 通过物理结合WNT 及其受体Frizzled 来抑制WNT 信号通路。Wnt 信号也受到分泌蛋白Dkk1 的抑制,Dkk1 是多基因家族的成员。Dkk1 通过与LRP5/6 结合并拮抗LRP5/6 来抑制Wnt 信号。最近有证据表明,跨膜蛋白Krm1 (Kremen-1)和Krm2 (Kremen-2)是高亲和力的Dkk1 受体,与Dkk1 在功能上协同阻断Wnt 信号。Krm2 与Dkk1 和LRP6 形成三元复合物,并诱导LRP6 迅速内吞和从质膜上的移除(参考文献6,7和8)。
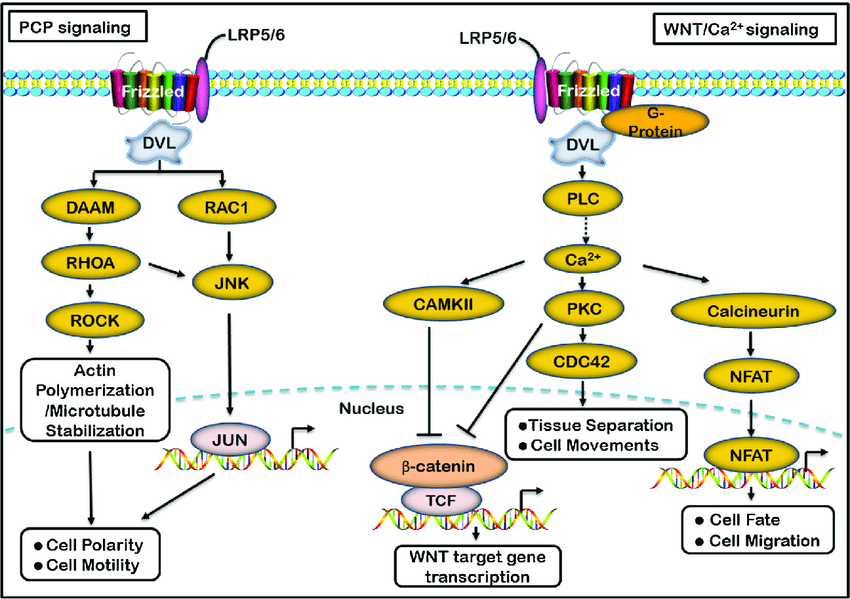
非经典Wnt信号通路,又称非经典Wnt-Frizzled信号通路,由Wnt/Ca 2+ 通路和Wnt/PCP通路两个细胞内信号级联通路组成。在WNT/Ca 2+ 途径中,WNT蛋白主要由WNT1、WNT5A和WNT11组成,与细胞表面的Frizzled跨膜受体结合,参与几个细胞过程,这些过程涉及刺激异三聚体G蛋白,从而进一步激活PLC(磷脂酶-C)。PLC导致细胞内Ca 2+ 释放增加,环鸟苷酸(CGMP)水平降低,CaMKII(Ca2+-钙调蛋白依赖性蛋白激酶-II)或Caln(钙调神经磷酸酶)和PKC(蛋白激酶-C)两种激酶被激活。这些过程可以刺激核因子NFAT和其他转录因子,如CREB(cAMP Response Element-Binding Protein-1)。因此,Wnt/Ca 2+ 通路很可能是G蛋白依赖的信号通路(参考文献3&9)。在Wnt/PCP途径中,Wnt蛋白与细胞表面的Frizzled跨膜受体结合,然后通过Dsh激活Rho/Rac小GTPase和JNK(Jun N端激酶),随后辅助细胞骨架组织和基因表达的调控。DSH通过Daam1连接到下游效应器Rho和ROCK(Rho关联激酶)。RAC由Dsh直接激活,Dsh通过分别激活MAP3Ks(丝裂原活化蛋白激酶)和MAP2Ks丝裂原活化蛋白激酶(MAP2Ks)进一步激活JNK。WNT靶基因NKD(Naked)的产物最近被鉴定为WNT信号的拮抗剂,它与Dsh结合,阻断Ctnn-Beta,但刺激JNK途径(参考文献3,10和11)。Wnt-Frizzled信号通路在神经系统发育的生物学中起着重要作用。特别是,Wnt-Frizzled信号通路参与了神经板与神经前体细胞的发育以及随后的神经管的前后延伸。最终,WNT-Frizzled信号通路导致大脑、脊髓的发育,以及感觉和运动神经元的众多亚群的延伸。WNT-Frizzled通路的功能障碍可能导致神经退行性疾病,如阿尔茨海默病和心力衰竭。通过进一步鉴别和靶向控制WNT-Frizzled信号通路的关键元件,可以更好地理解WNT-Frizzled信号通路的生物学潜能,从而开发针对神经退行性疾病和血管疾病的新的治疗方案(参考文献12&13)。 往期信号通路介绍汇总(点击标题直达文章) 参考资料: 1. Li F, Chong ZZ, Maiese K. Windingthrough the WNT pathway during cellular development and demise. HistolHistopathol. 2006 Jan;21(1):103-24. PubMed ID: 16267791 2. Katoh M, Katoh M. Comparative genomicson Wnt5a and Wnt5b genes. Int J Mol Med. 2005 Apr;15(4):749-53. PubMed ID:15754042 3. Li F, Chong ZZ, Maiese K. Vital elementsof the Wnt-Frizzled signaling pathway in the nervous system. Curr NeurovascRes. 2005 Oct;2(4):331-40. PubMed ID: 16181124 4. Urano T. Wnt-beta-catenin signaling inbone metabolism. Clin Calcium. 2006 Jan;16(1):54-60. PubMed ID: 16397351 5. Souaze F, Viardot-Foucault V, Roullet N,Toy-Miou-Leong M, Gompel A, Bruyneel E, Comperat E, Faux MC, Mareel M, RosteneW, Flejou JF, Gespach C, Forgez P. Neurotensin receptor 1 gene activation by theTcf/{beta}-catenin pathway is an early event in human colonic adenomas. Carcinogenesis.2005 Nov 19; PubMed ID: 16299383 6. Blavier L, Lazaryev A, Dorey F,Shackleford GM, DeClerck YA. Matrix metalloproteinases play an active role in Wnt1-inducedmammary tumorigenesis. Cancer Res. 2006 Mar 1;66(5):2691-9. PubMed ID: 16510589 7. Kikuchi A, Kishida S, Yamamoto H. Regulationof Wnt signaling by protein-protein interaction and post-translationalmodifications. Exp Mol Med. 2006 Feb 28;38(1):1-10. PubMed ID: 16520547 8. Mao B, Wu W, Davidson G, Marhold J, LiM, Mechler BM, Delius H, Hoppe D, Stannek P, Walter C, Glinka A, Niehrs C. Kremenproteins are Dickkopf receptors that regulate Wnt/beta-catenin signalling. Nature.2002 Jun 6;417(6889):664-7. Epub 2002 May 26. PubMed ID: 12050670 9. Kohn AD, Moon RT. Wnt and calciumsignaling: beta-catenin-independent pathways. Cell Calcium. 2005Sep-Oct;38(3-4):439-46. PubMed ID: 16099039 10. Goldstein B, Takeshita H, Mizumoto K,Sawa H. Wnt signals can function as positional cues in establishing cellpolarity. Dev Cell. 2006 Mar;10(3):391-6. PubMed ID: 16516841 11. Katoh M. WNT/PCP signaling pathway andhuman cancer. Oncol Rep. 2005 Dec;14(6):1583-8. PubMed ID: 16273260 12. Timm A, Grosschedl R. Wnt signaling inlymphopoiesis. Curr Top Microbiol Immunol. 2005;290:225-52. PubMed ID: 16480045 13. Janssens N, Janicot M, Perera T. TheWnt-dependent signaling pathways as target in oncology drug discovery. InvestNew Drugs. 2006 Jan 28; PubMed ID: 16444610 科研小助手 官方QQ群 :93646661转载、投稿、商务合作amateur_1988 转载、投稿、商务合作amateur_1988返回搜狐,查看更多 |
【本文地址】
今日新闻 |
推荐新闻 |